

FoxP3+ regulatory T cells promote influenza-specific Tfh responses by controlling IL-2 availability
- PMID: 24633065
- PMCID: PMC4013682
- DOI: 10.1038/ncomms4495
FoxP3+ regulatory T cells promote influenza-specific Tfh responses by controlling IL-2 availability
Abstract
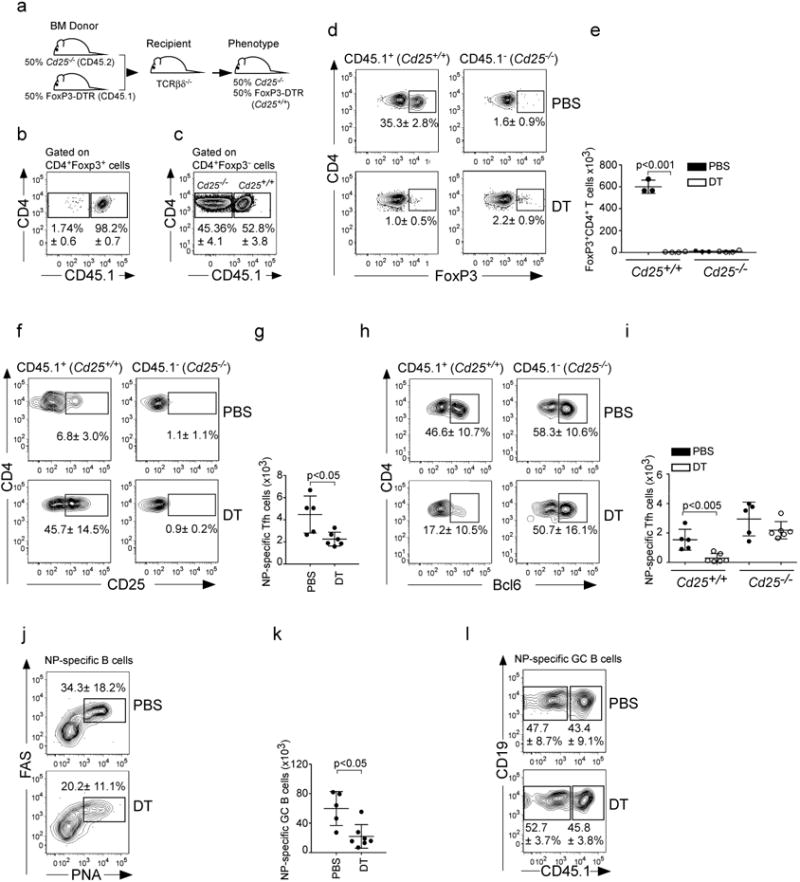
Here, we test the role of FoxP3(+) regulatory T cells (Tregs) in controlling T follicular helper (Tfh) and germinal centre (GC) B-cell responses to influenza. In contrast to the idea that Tregs suppress T-cell responses, we find that Treg depletion severely reduces the Tfh cell response to influenza virus. Furthermore, Treg depletion prevents the accumulation of influenza-specific GCs. These effects are not due to alterations in TGFβ availability or a precursor-progeny relationship between Tregs and Tfh cells, but are instead mediated by increased availability of IL-2, which suppresses the differentiation of Tfh cells and as a consequence, compromises the GC B response. Thus, Tregs promote influenza-specific GC responses by preventing excessive IL-2 signalling, which suppresses Tfh cell differentiation.
Figures






Comment in
-
Regulatory T cells: promoting B cell responses to influenza virus.Nat Rev Immunol. 2014 May;14(5):283. doi: 10.1038/nri3662. Epub 2014 Apr 7. Nat Rev Immunol. 2014. PMID: 24705704 No abstract available.
References
-
- Goodnow CC, Vinuesa CG, Randall KL, Mackay F, Brink R. Control systems and decision making for antibody production. Nature immunology. 2010;11:681–688. - PubMed
-
- Vinuesa CG, Linterman MA, Goodnow CC, Randall KL. T cells and follicular dendritic cells in germinal center B-cell formation and selection. Immunological reviews. 2010;237:72–89. - PubMed
-
- Crotty S. Follicular helper CD4 T cells (TFH) Annual review of immunology. 2011;29:621–663. - PubMed
-
- Yu D, Vinuesa CG. The elusive identity of T follicular helper cells. Trends in immunology. 2010;31:377–383. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
