

Association between foldability and aggregation propensity in small disulfide-rich proteins
- PMID: 24635049
- PMCID: PMC4076991
- DOI: 10.1089/ars.2013.5543
Association between foldability and aggregation propensity in small disulfide-rich proteins
Abstract
Aims: Disulfide-rich domains (DRDs) are small proteins whose native structure is stabilized by the presence of covalent disulfide bonds. These domains are versatile and can perform a wide range of functions. Many of these domains readily unfold on disulfide bond reduction, suggesting that in the absence of covalent bonding they might display significant disorder.
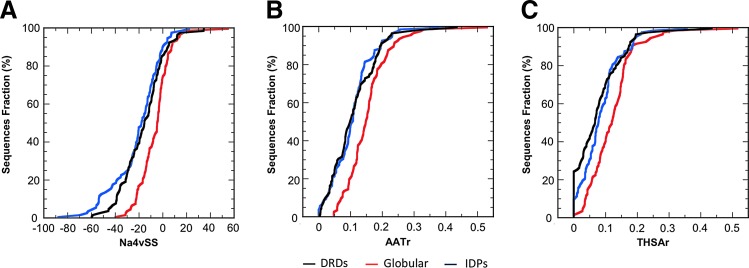
Results: Here, we analyzed the degree of disorder in 97 domains representative of the different DRDs families and demonstrate that, in terms of sequence, many of them can be classified as intrinsically disordered proteins (IDPs) or contain predicted disordered regions. The analysis of the aggregation propensity of these domains indicates that, similar to IDPs, their sequences are more soluble and have less aggregating regions than those of other globular domains, suggesting that they might have evolved to avoid aggregation after protein synthesis and before they can attain its compact and covalently linked native structure.
Innovation and conclusion: DRDs, which resemble IDPs in the reduced state and become globular when their disulfide bonds are formed, illustrate the link between protein folding and aggregation propensities and how these two properties cannot be easily dissociated, determining the main traits of the folding routes followed by these small proteins to attain their native oxidized states.
Figures








References
-
- Arias-Moreno X, Arolas JL, Aviles FX, Sancho J, and Ventura S. Scrambled isomers as key intermediates in the oxidative folding of ligand binding module 5 of the low density lipoprotein receptor. J Biol Chem 283: 13627–13637, 2008 - PubMed
-
- Arolas JL, Aviles FX, Chang JY, and Ventura S. Folding of small disulfide-rich proteins: clarifying the puzzle. Trends Biochem Sci 31: 292–301, 2006 - PubMed
-
- Arolas JL, Bronsoms S, Aviles FX, Ventura S, and Sommerhoff CP. Oxidative folding of leech-derived tryptase inhibitor via native disulfide-bonded intermediates. Antioxid Redox Signal 10: 77–85, 2008 - PubMed
-
- Arolas JL, Bronsoms S, Lorenzo J, Aviles FX, Chang JY, and Ventura S. Role of kinetic intermediates in the folding of leech carboxypeptidase inhibitor. J Biol Chem 279: 37261–37270, 2004 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
