

Chlorophyllide a oxidoreductase works as one of the divinyl reductases specifically involved in bacteriochlorophyll a biosynthesis
- PMID: 24637023
- PMCID: PMC4007461
- DOI: 10.1074/jbc.M113.546739
Chlorophyllide a oxidoreductase works as one of the divinyl reductases specifically involved in bacteriochlorophyll a biosynthesis
Abstract
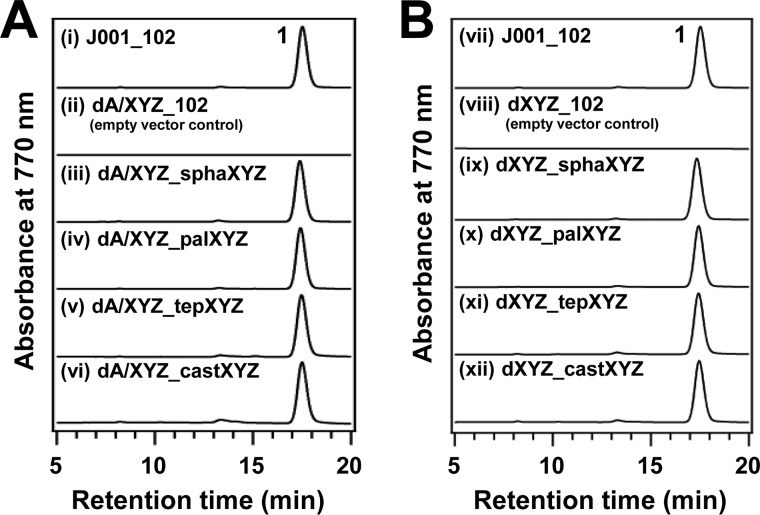
Bacteriochlorophyll a is widely distributed among anoxygenic photosynthetic bacteria. In bacteriochlorophyll a biosynthesis, the reduction of the C8 vinyl group in 8-vinyl-chlorophyllide a is catalyzed to produce chlorophyllide a by an 8-vinyl reductase called divinyl reductase (DVR), which has been classified into two types, BciA and BciB. However, previous studies demonstrated that mutants lacking the DVR still synthesize normal bacteriochlorophyll a with the C8 ethyl group and suggested the existence of an unknown "third" DVR. Meanwhile, we recently observed that chlorophyllide a oxidoreductase (COR) of a purple bacterium happened to show the 8-vinyl reduction of 8-vinyl-chlorophyllide a in vitro. In this study, we made a double mutant lacking BciA and COR of the purple bacterium Rhodobacter sphaeroides in order to investigate whether the mutant still produces pigments with the C8 ethyl group or if COR actually works as the third DVR. The single mutant deleting BciA or COR showed production of the C8 ethyl group pigments, whereas the double mutant accumulated 8-vinyl-chlorophyllide, indicating that there was no enzyme other than BciA and COR functioning as the unknown third DVR in Rhodobacter sphaeroides (note that this bacterium has no bciB gene). Moreover, some COR genes derived from other groups of anoxygenic photosynthetic bacteria were introduced into the double mutant, and all of the complementary strains produced normal bacteriochlorophyll a. This observation indicated that COR of these bacteria performs two functions, reductions of the C8 vinyl group and the C7=C8 double bond, and that such an activity is probably conserved in the widely ranging groups.
Keywords: BciA; BciB; DVR; Evolution; Microbiology; Photosynthesis; Photosynthetic Bacteria; Photosynthetic Pigments; Purple Bacteria; Reductase.
Figures




References
-
- Tanaka R., Tanaka A. (2007) Tetrapyrrole biosynthesis in higher plants. Annu. Rev. Plant Biol. 58, 321–346 - PubMed
-
- Miyashita H., Ikemoto H., Kurano N., Adachi K., Chihara M., Miyachi S. (1996) Chlorophyll d as a major pigment. Nature 10.1038/383402a0 - DOI
-
- Chen M., Schliep M., Willows R. D., Cai Z.-L., Neilan B. A., Scheer H. (2010) A red-shifted chlorophyll. Science 329, 1318–1319 - PubMed
-
- Bollivar D. W., Suzuki J. Y., Beatty J. T., Dobrowolski J. M., Bauer C. E. (1994) Directed mutational analysis of bacteriochlorophyll a biosynthesis in Rhodobacter capsulatus. J. Mol. Biol. 237, 622–640 - PubMed
-
- Chew A. G., Bryant D. A. (2007) Chlorophyll biosynthesis in bacteria: the origins of structural and functional diversity. Annu. Rev. Microbiol. 61, 113–129 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
