

Review
doi: 10.1085/jgp.201411164.
Epub 2014 Mar 17.
Structural diversity of ABC transporters
Affiliations
- PMID: 24638992
- PMCID: PMC3971661
- DOI: 10.1085/jgp.201411164
Item in Clipboard
Review
Structural diversity of ABC transporters
J Gen Physiol.
2014 Apr.
Abstract
ATP-binding cassette (ABC) transporters form a large superfamily of ATP-dependent protein complexes that mediate transport of a vast array of substrates across membranes. The 14 currently available structures of ABC transporters have greatly advanced insight into the transport mechanism and revealed a tremendous structural diversity. Whereas the domains that hydrolyze ATP are structurally related in all ABC transporters, the membrane-embedded domains, where the substrates are translocated, adopt four different unrelated folds. Here, we review the structural characteristics of ABC transporters and discuss the implications of this structural diversity for mechanistic diversity.
Figures
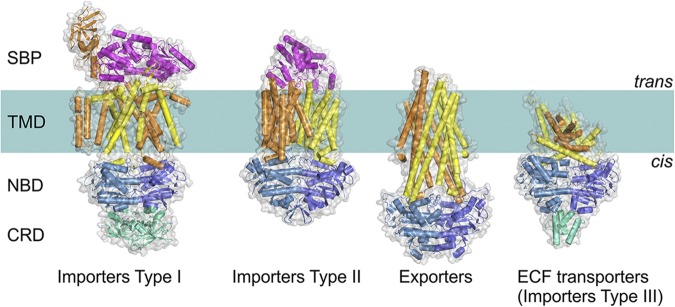
Four distinct folds of ABC transporters. All share a similar general architecture: two NBDs (blue and sky blue) are attached to two TMDs (orange and yellow). In some transporters, additional domains are present (green), which often have a regulatory function (C-terminal regulatory domain [CRD]). In Type I and II importers, the transported compounds are delivered to TMDs by SBPs (or SBDs; magenta) located in periplasm (Gram-negative bacteria) or external space (Gram-positive bacteria and Archaea). ECF, energy coupling factor.
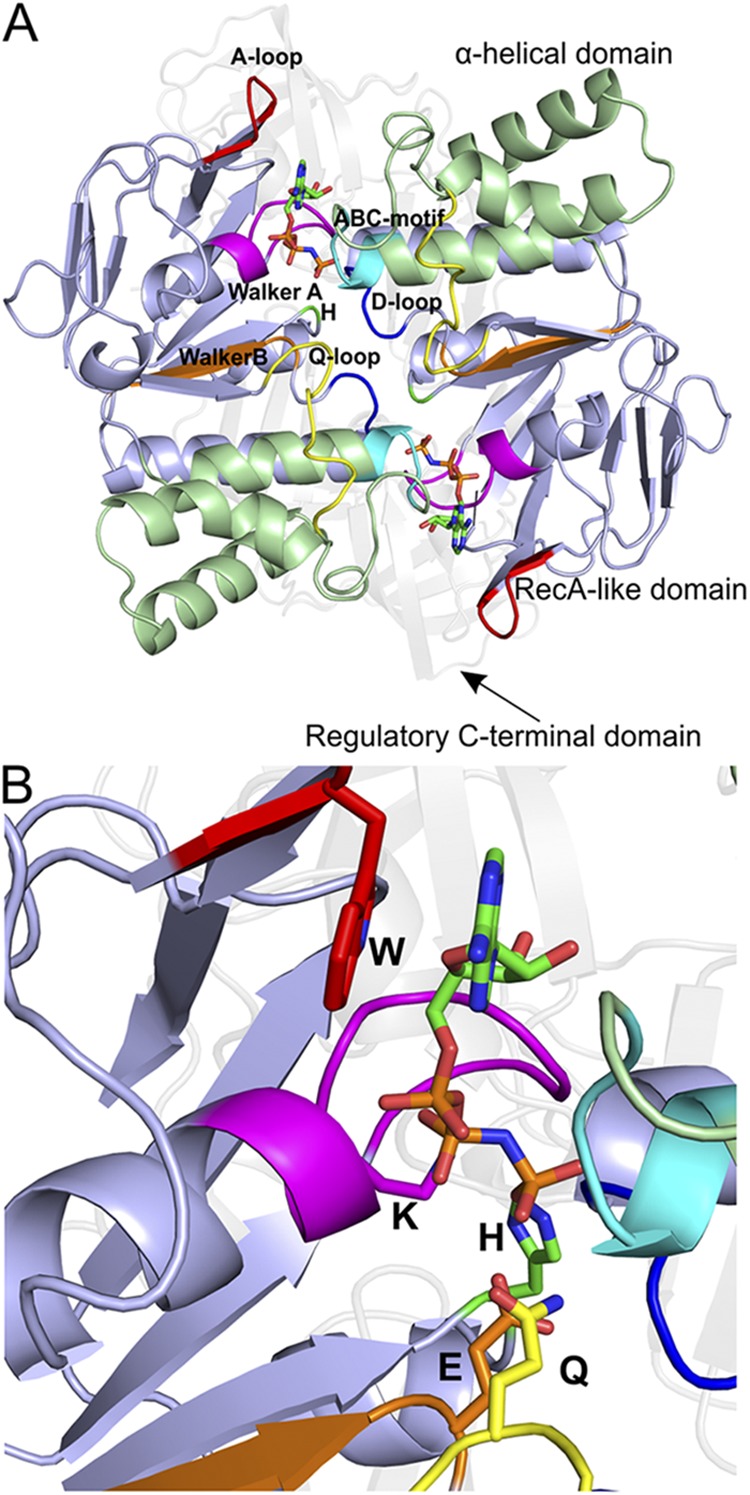
The structure of the NBDs, as exemplified by the MalK dimer of the maltose transporter MalEFGK2 (Protein Data Bank accession no. 3RLF). (A) View along an axis perpendicular to the membrane plane from the trans-side onto the NBDs (The TMDs and SBP have been removed for clarity). Domains and highly conserved sequence motifs are color-coded: green, α-helical domain; light blue, RecA-like domain; faded gray, regulatory C-terminal domain; red, A-loop; magenta, Walker A; orange, Walker B; blue, D-loop; green, H-loop; cyan, ABC motif; yellow, Q-loop. The ATP analogue AMP-PNP is shown in sticks. (B) A closer look onto the nucleotide-binding site. The key amino acids are indicated (see NBD for details).
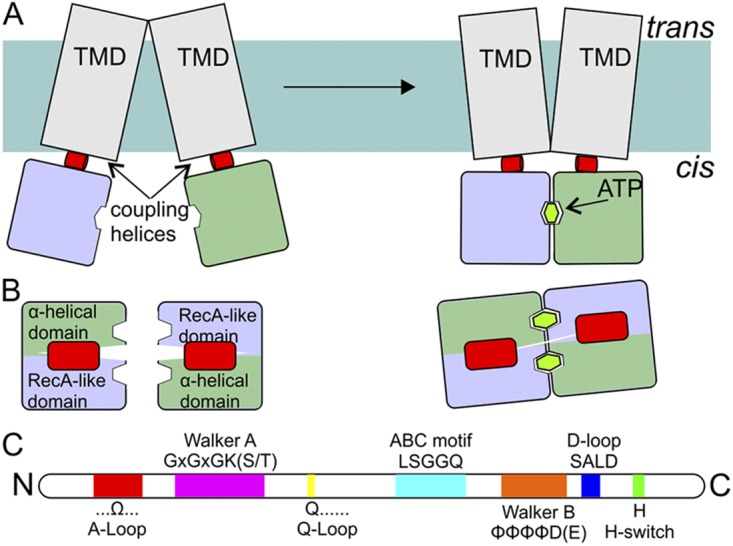
Schematic representation of NBDs and coupling helices. (A) Side view (from the membrane plane) of an ABC transporter. NBDs (blue and green; colors of the domains are as in Fig. 2) are attached to the TMDs (gray) via so-called coupling helices (red) present in loops of the TMDs. ATP binding and hydrolysis cause rearrangements in the NBDs, which are propagated to the TMDs via the coupling helices. (B) Top view (along an axis perpendicular to the membrane) of the NBDs and the coupling helices from the TMDs. (C) The relative positions of sequence motifs in NBDs (see also Fig. 2).
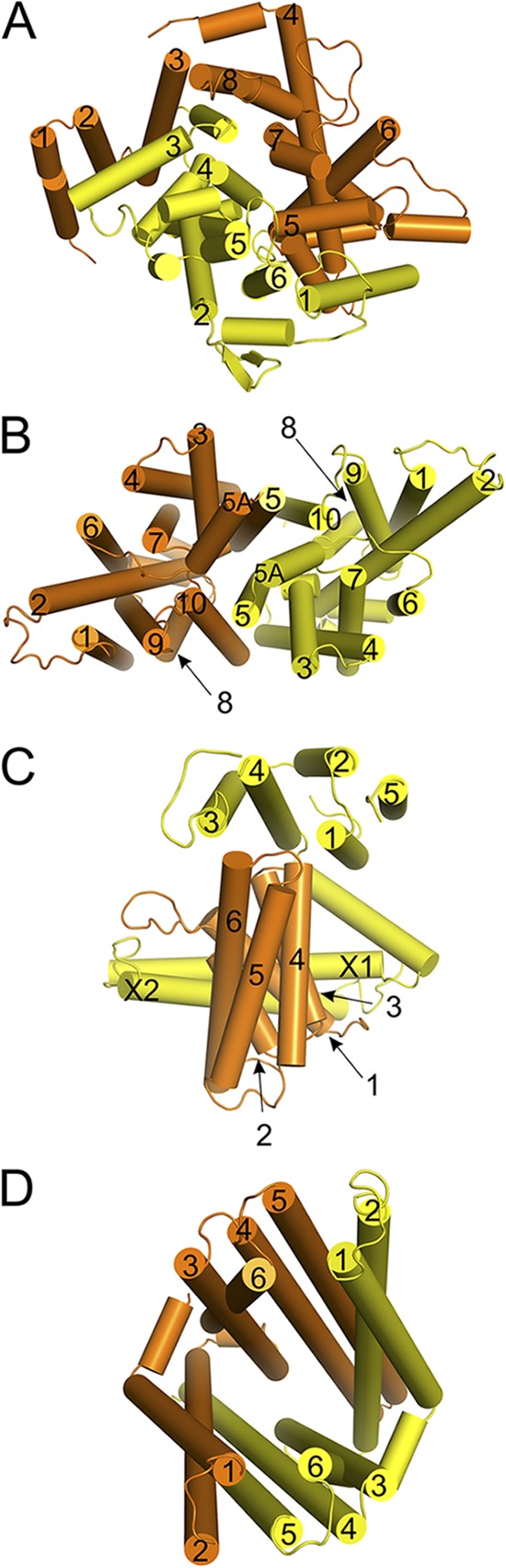
Arrangement of the membrane helices in ABC transporters. Viewpoints are from the outside (trans-side) along an axis perpendicular to the membrane plane. (A) The MalF and MalG subunits of the maltose transporter MalEFGK2, (B) the BtuC dimer of the vitamin B12 transporter BtuC2D2F, (C) the EcfT subunit (yellow) and the S-component from the ECF-HmpT transporter, and (D) the membrane domains of the TM287 and TM288 subunits of the exporter from T. maritima. TM helices are numbered according to their occurrence in the sequence, with the one located most closely to the N terminus numbered as 1. The two coupling helices in the EcfT subunit are labeled X1 and X2.
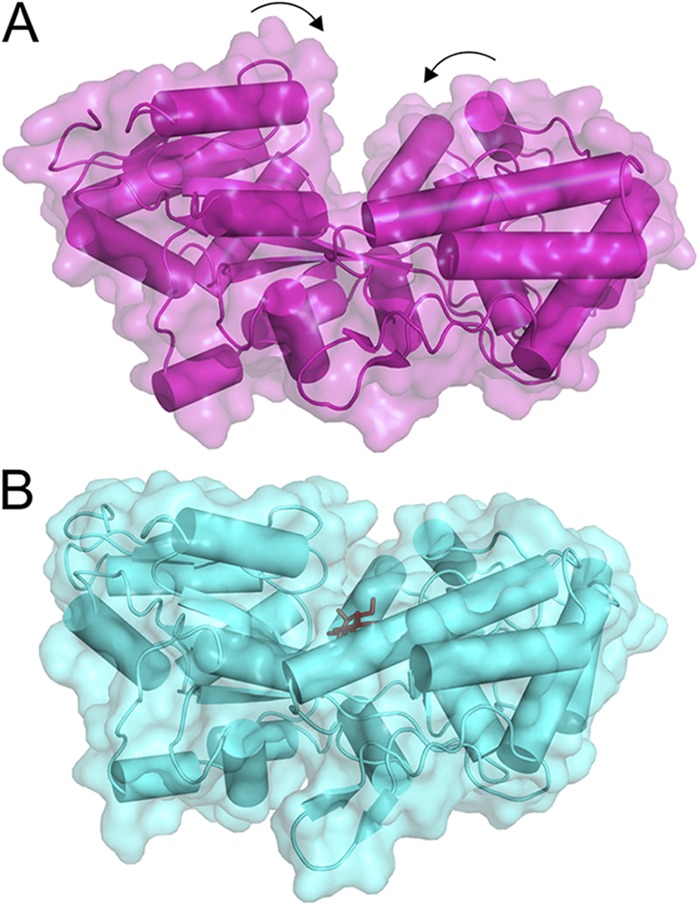
Rearrangements in MBP upon the substrate binding. (A) In the substrate-free form (Protein Data Bank accession no. 1ANF), the cavity between two protein lobes connected by the hinge is accessible. (B) Upon the binding of substrate (dark sticks; Protein Data Bank accession no. 1EZ9), the cavity becomes occluded.
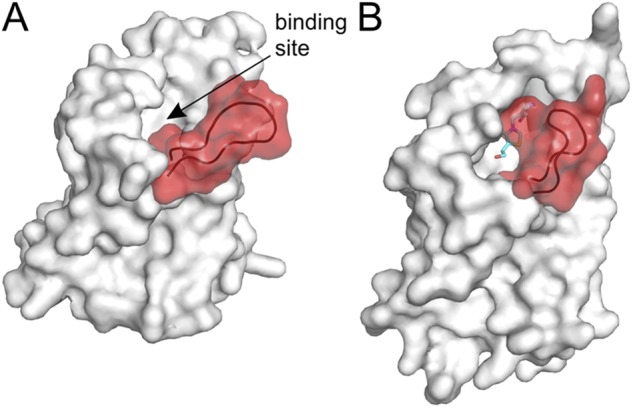
Structural changes in the S-component of ECF transporters (substrate-binding TM protein of ECF transporters). The loop between TM helices 1 and 2 is highlighted in red. (A) S-component from the ECF–HmpT complex (Protein Data Bank accession no. 4HZU) in the substrate-free form. The shape of the binding pocket is visible, indicated by the arrow. (B) Thiamin-specific ThiT (Protein Data Bank accession no. 3RLB) in the substrate-bound state. The loop between TM helices 1 and 2 closes the binding pocket. The substrate thiamin is shown as sticks.
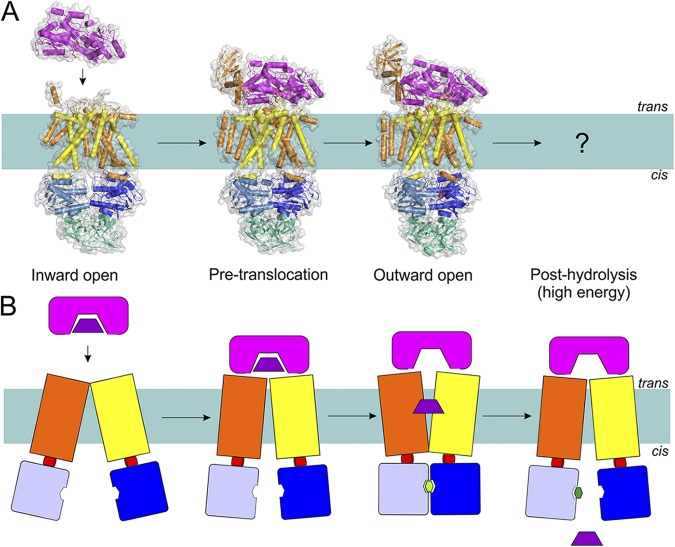
The transport mechanism of Type I importers (exemplified by MalEFGK2) based on the available structures (A) and in schematic representation (B). Coloring is as in Fig. 1. Structures have been determined for the inward-facing, pre-translocation, and outward-facing conformations (Protein Data Bank accession nos.: 4JBW, 4KHZ, and 4KI0; see Table 1 for details of all available structures of ABC transporters). Substrate-loaded MalE docks onto the resting MalFGK2 transporter. This brings the NBDs closer to each other, allows ATP binding, and consequently causes MalK closure and TMD reorientation (outward-facing), leading to MBP opening. The released substrate diffuses toward the binding site in TMD, followed by ATP hydrolysis and resetting of the transporter into the inward-facing state.
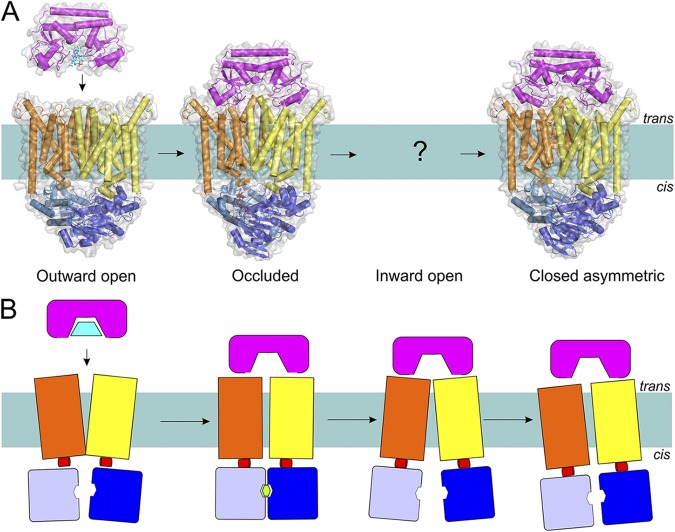
The transport mechanism of Type II importers (exemplified by the BtuC2D2F transporter) based on the available structures (A) and in schematic representation (B). Coloring is as in Fig. 1. Structures have been determined for an outward-open, occluded nucleotide-bound, and closed ATP-free asymmetric transporter (Protein Data Bank accession nos.: 1L7V, 4DBL, and 2QI9; see Table 1 for details of all available structures of ABC transporters). Substrate-loaded BtuF docks onto the resting BtuC2D2 transporter in the outward conformation. ATP binding and concomitant rearrangements in the TMDs lead to the trapping of the substrate between TMDs (substrate was not visible in the crystal structure). The hydrolysis of ATP causes opening of the cytoplasmic gate and allows substrate release, after which the gate is asymmetrically closed, possibly to prevent the leakage of small molecules.
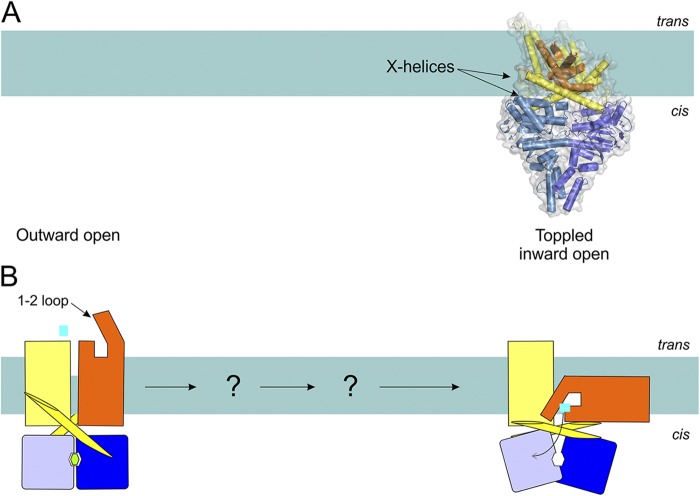
Possible transport mechanism of ECF transporters (Type III importers) based on the available structures (A) and in schematic representation (B). Structures have been determined of the nucleotide- and substrate-free transporter (ECF-HmpT, Protein Data Bank accession no. 4HZU, is shown here; see Table 1 for details of all available structures of ABC transporters). The S-component (orange) is predicted to rotate in the membrane (“topple over”) to bring the substrate from the trans-side to the cis-side.
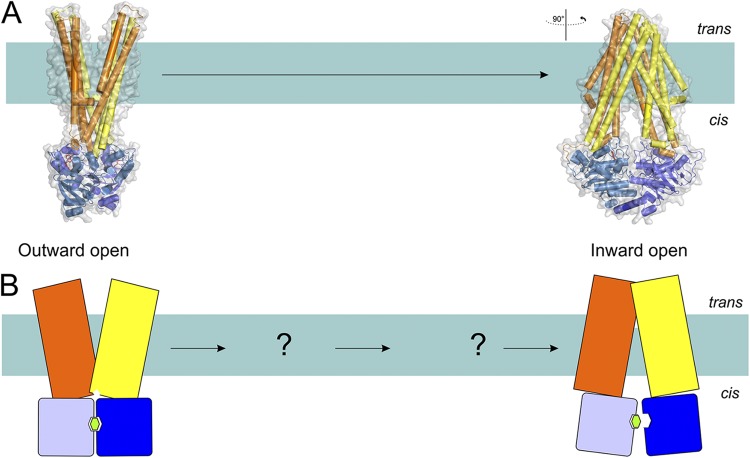
The transport mechanism of exporters (exemplified by the structures of Sav18662 and TM287/288) based on the available structures (A) and in schematic representation (B). Coloring is as in Fig. 1. Structures of outward-open and nucleotide-bound inward-facing transporter are shown (Protein Data Bank accession nos. 2HYD and 3QF4; see Table 1 for details of all available structures of ABC transporters). The hydrolysis of ATP in the outward-open conformation results in the reorientation of TMDs, leading to the inward-open conformation where substrate and ATP binding take place. This in turn orients the transporter back into the outward-open conformation, and the substrate is expelled.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
