

Canine lymphoma as a comparative model for human non-Hodgkin lymphoma: recent progress and applications
- PMID: 24642290
- PMCID: PMC4994713
- DOI: 10.1016/j.vetimm.2014.02.016
Canine lymphoma as a comparative model for human non-Hodgkin lymphoma: recent progress and applications
Abstract
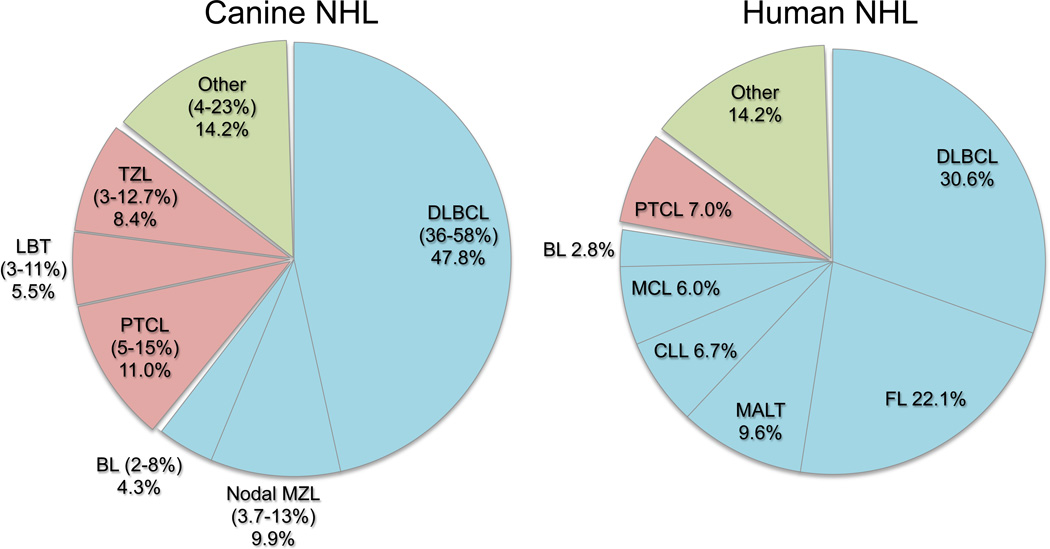
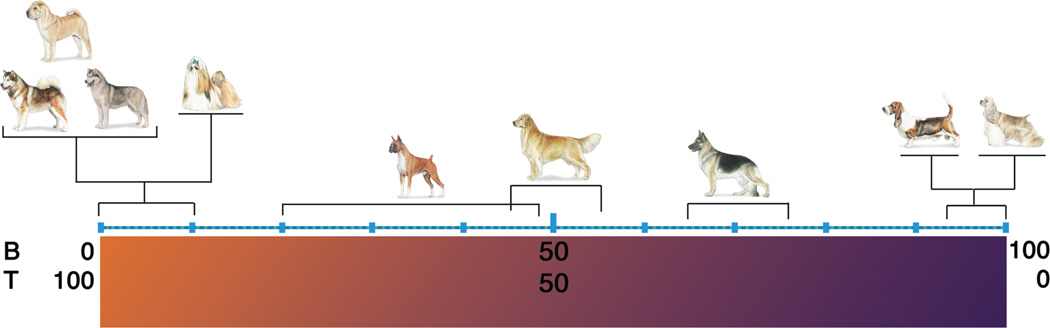
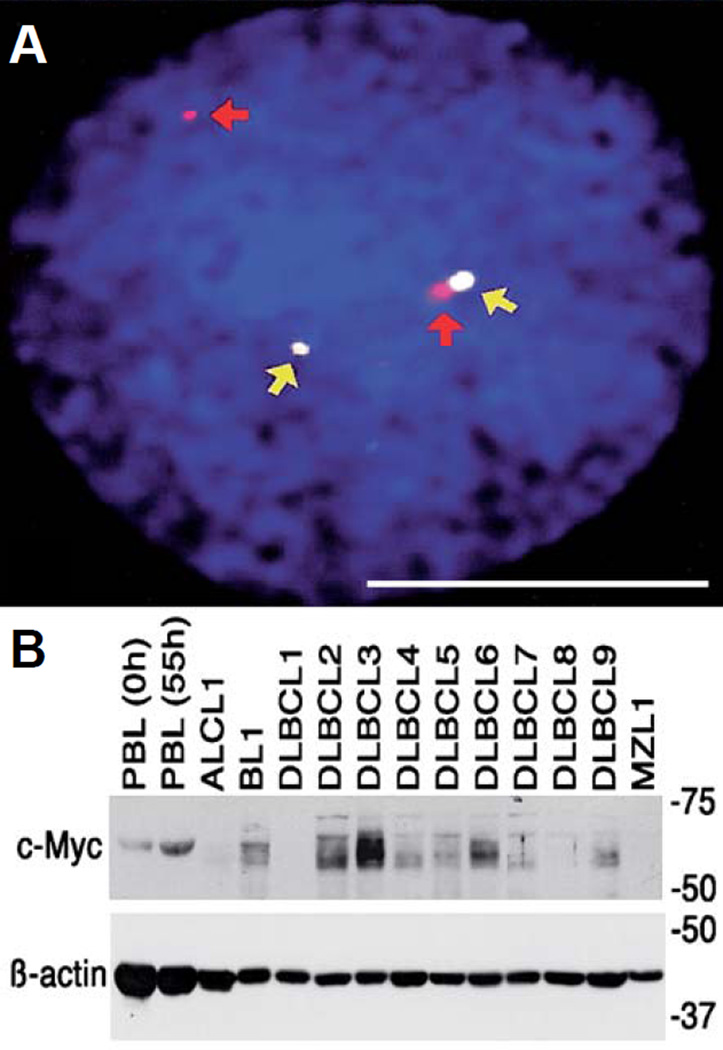
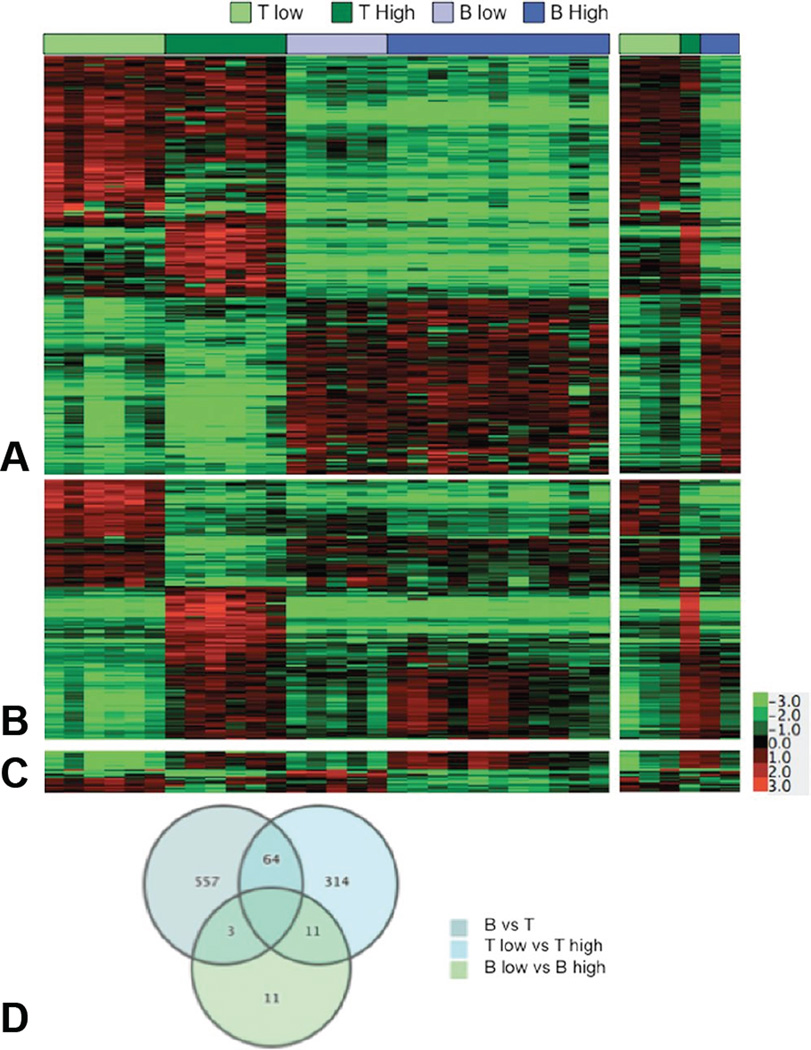
The term "lymphoma" describes a heterogeneous group of disorders involving monoclonal proliferation of malignant lymphocytes. As a group, lymphomas are among the most common tumors of dogs. Yet our enumeration and understanding of the many subtypes of lymphoma have been relatively slow, perhaps in part because for many years lymphoma was treated as a singular entity rather than a group of distinct diseases. The recognition that the full spectrum of lymphoid malignancies seen in humans also occurs in dogs, and that these tumors retain not only morphologic similarities and biological behavior but also synonymous driver molecular abnormalities, sets an ideal stage for dual-purpose research that can accelerate progress for these diseases in both species. Specifically, dogs represent exceptional models for defining causality, understanding progression, and developing new treatments for lymphoma in comparatively brief windows of time. Unique advantages of canine models include (1) spontaneous disease occurring without an isogenic background or genetic engineering; (2) chronology of disease adapted to lifespan, (3) shared environment and societal status that allows dogs to be treated as "patients," while at the same time being able to ethically explore translational innovations that are not possible in human subjects; and (4) organization of dogs into breeds with relatively homogeneous genetic backgrounds and distinct predisposition for lymphomas. Here, we will review recent studies describing intrinsic and extrinsic factors that contribute to the pathogenesis of canine and human lymphomas, as well as newly developed tools that will enhance the fidelity of these models to improve diagnosis and develop new treatments.
Copyright © 2014 Elsevier B.V. All rights reserved.
Figures
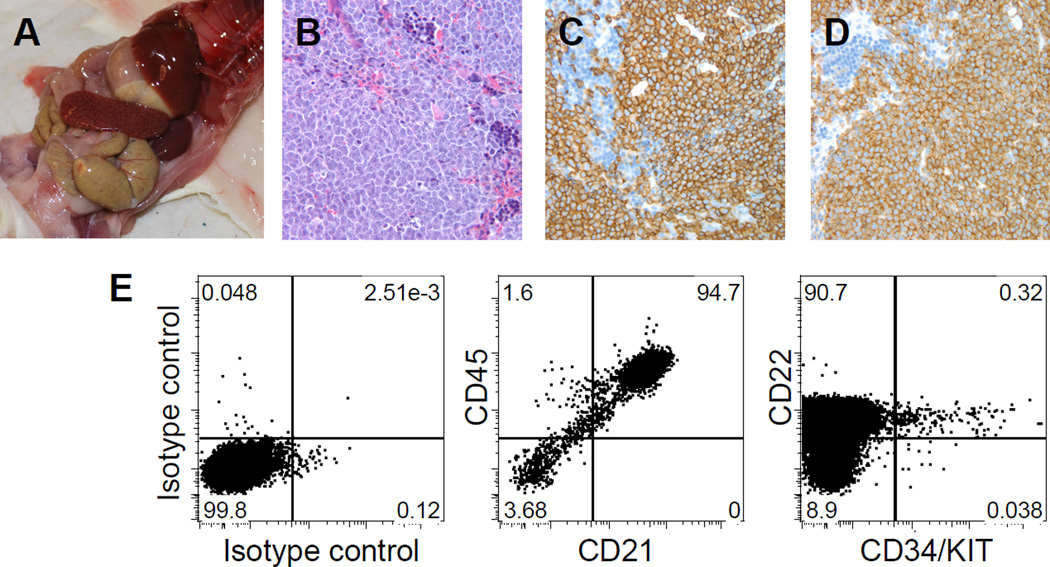
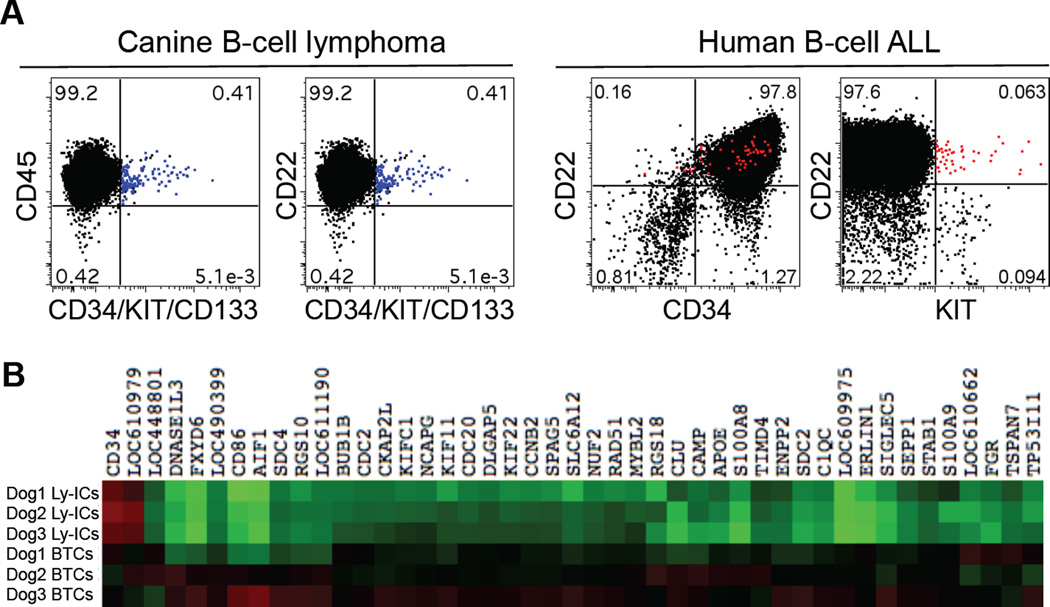
References
-
- Alizadeh AA, Eisen MB, Davis RE, Ma C, Lossos IS, Rosenwald A, Boldrick JC, Sabet H, Tran T, Yu X, Powell JI, Yang L, Marti GE, Moore T, Hudson J, Lu L, Lewis DB, Tibshirani R, Sherlock G, Chan WC, Greiner TC, Weisenburger DD, Armitage JO, Warnke R, Levy R, Wilson W, Grever MR, Byrd JC, Botstein D, Brown PO, Staudt LM. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature. 2000;403:503–511. - PubMed
-
- Alvarez CE, Akey JM. Copy number variation in the domestic dog. Mammalian Genome. 2012;23:144–163. - PubMed
-
- Andersen NS, Larsen JK, Christiansen J, Pedersen LB, Christophersen NS, Geisler CH, Jurlander J. Soluble CD40 ligand induces selective proliferation of lymphoma cells in primary mantle cell lymphoma cell cultures. Blood. 2000;96:2219–2225. - PubMed
-
- Anon A clinical evaluation of the International Lymphoma Study Group classification of non-Hodgkin's lymphoma. The non-Hodgkin's lymphoma classification project. Blood. 1997;89:3909–3918. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
