

Long term functional plasticity of sensory inputs mediated by olfactory learning
- PMID: 24642413
- PMCID: PMC3953949
- DOI: 10.7554/eLife.02109
Long term functional plasticity of sensory inputs mediated by olfactory learning
Abstract
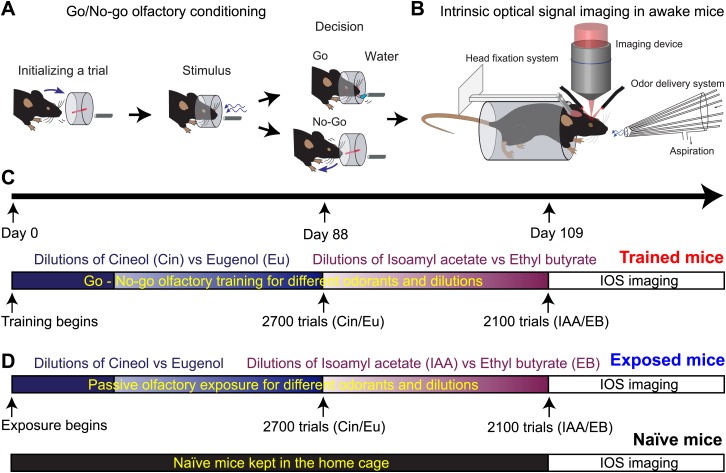
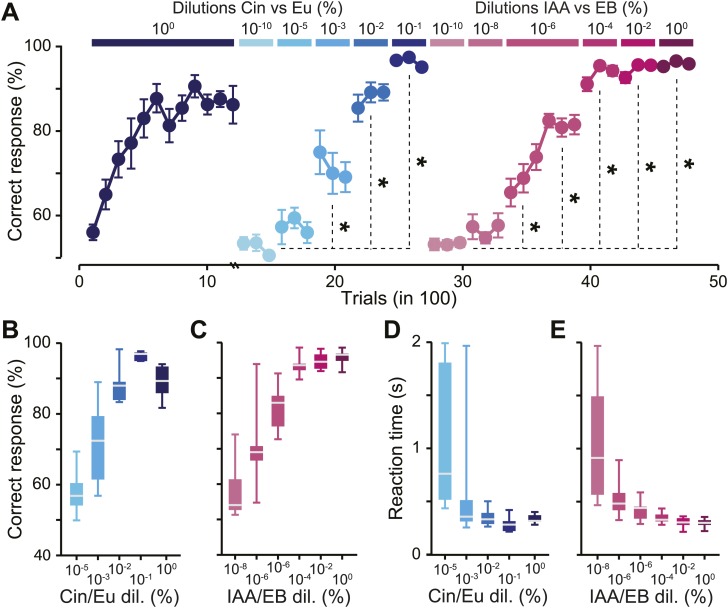
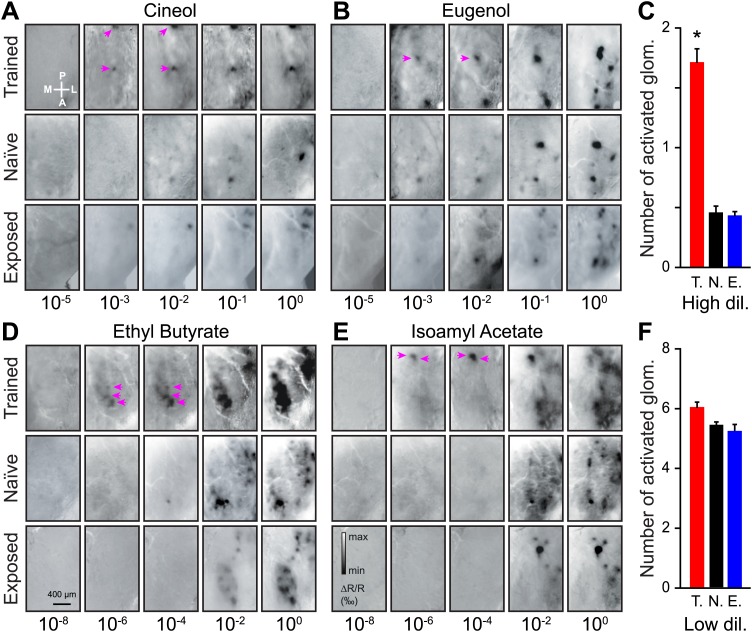
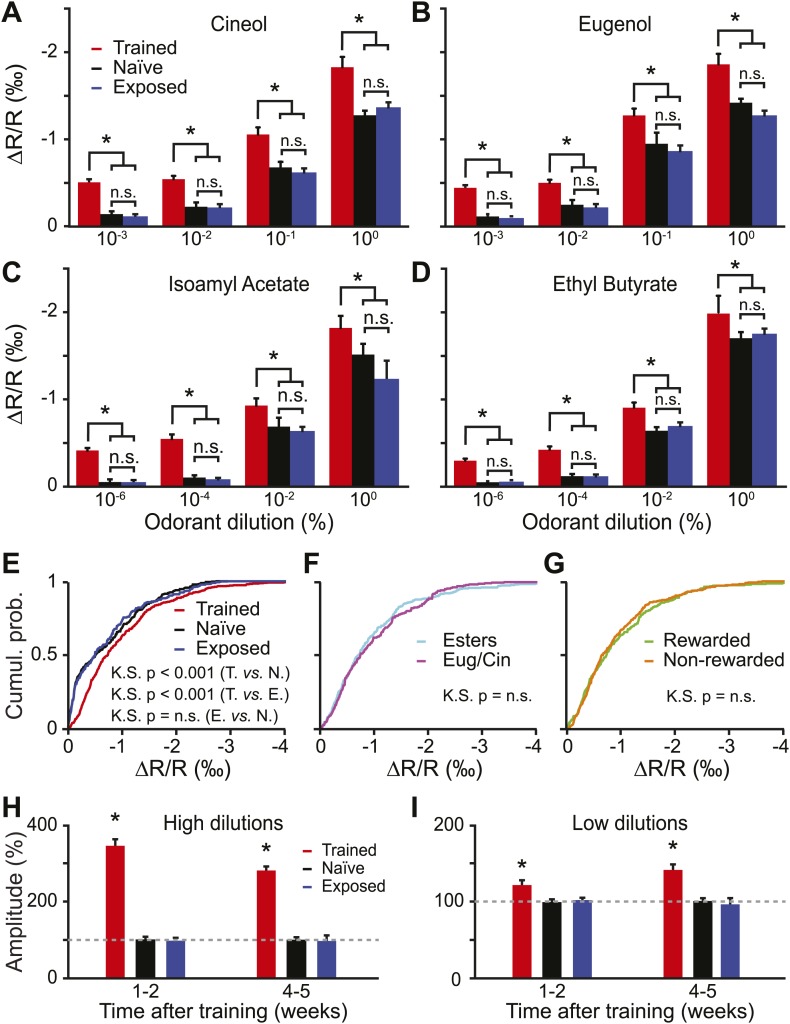
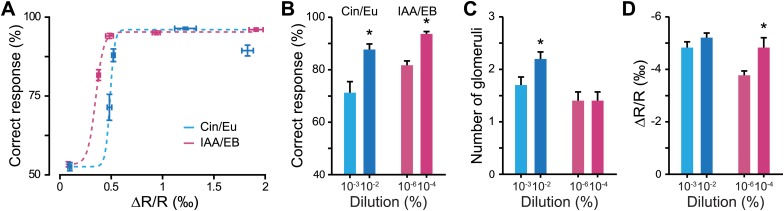
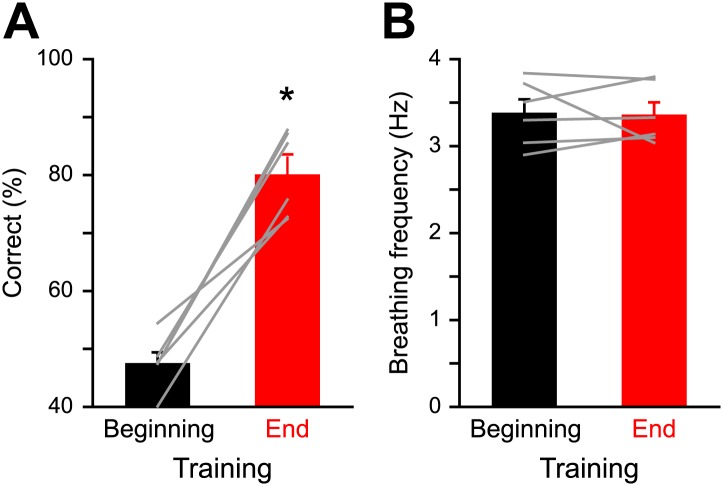
Sensory inputs are remarkably organized along all sensory pathways. While sensory representations are known to undergo plasticity at the higher levels of sensory pathways following peripheral lesions or sensory experience, less is known about the functional plasticity of peripheral inputs induced by learning. We addressed this question in the adult mouse olfactory system by combining odor discrimination studies with functional imaging of sensory input activity in awake mice. Here we show that associative learning, but not passive odor exposure, potentiates the strength of sensory inputs up to several weeks after the end of training. We conclude that experience-dependent plasticity can occur in the periphery of adult mouse olfactory system, which should improve odor detection and contribute towards accurate and fast odor discriminations. DOI: http://dx.doi.org/10.7554/eLife.02109.001.
Keywords: behavior; imaging; sensory perception.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
