

Subcellular localization of ENS-1/ERNI in chick embryonic stem cells
- PMID: 24643087
- PMCID: PMC3958431
- DOI: 10.1371/journal.pone.0092039
Subcellular localization of ENS-1/ERNI in chick embryonic stem cells
Abstract
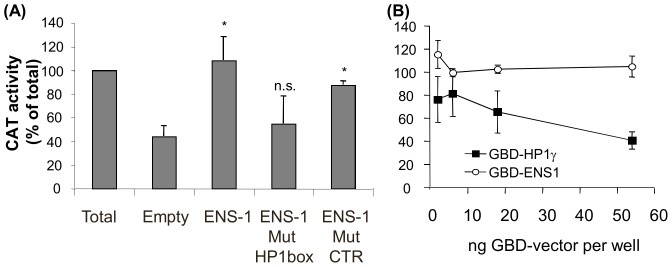
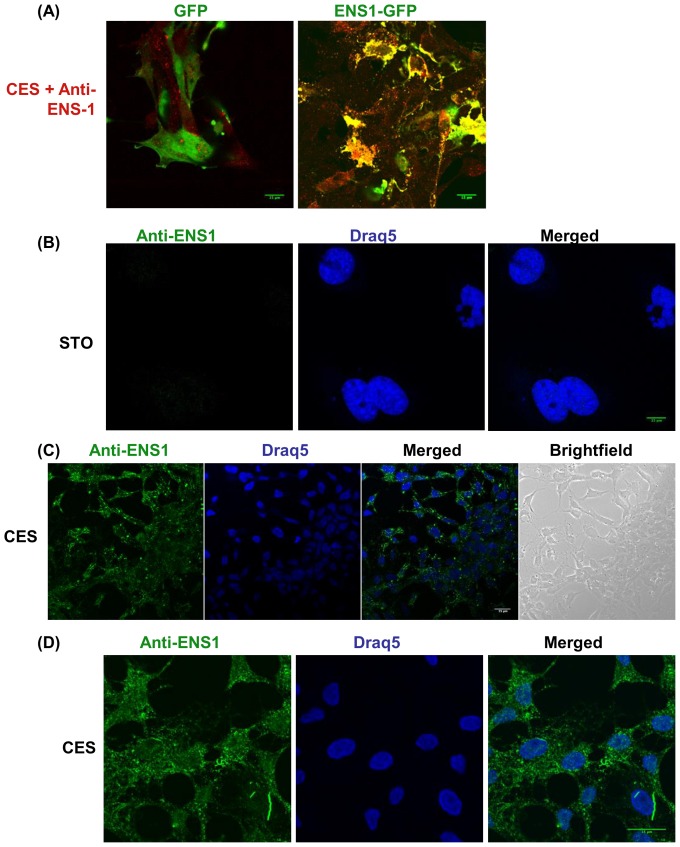
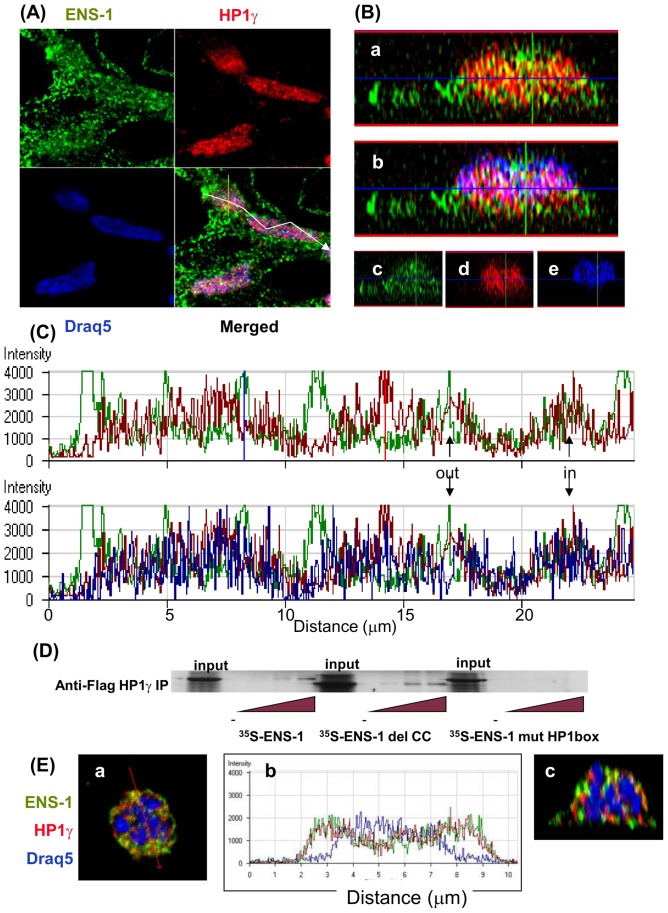
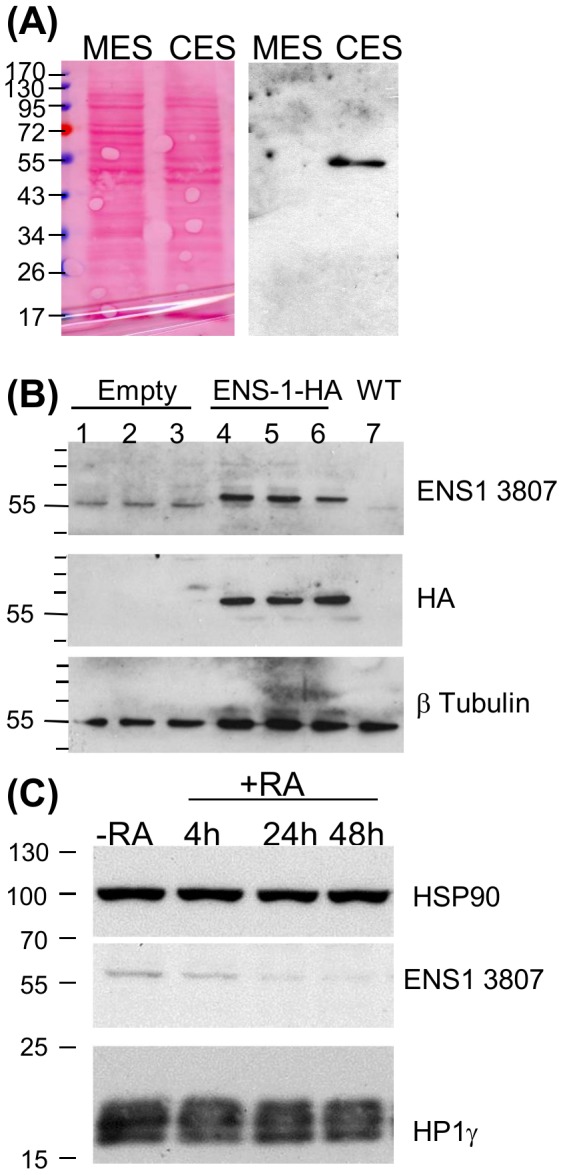
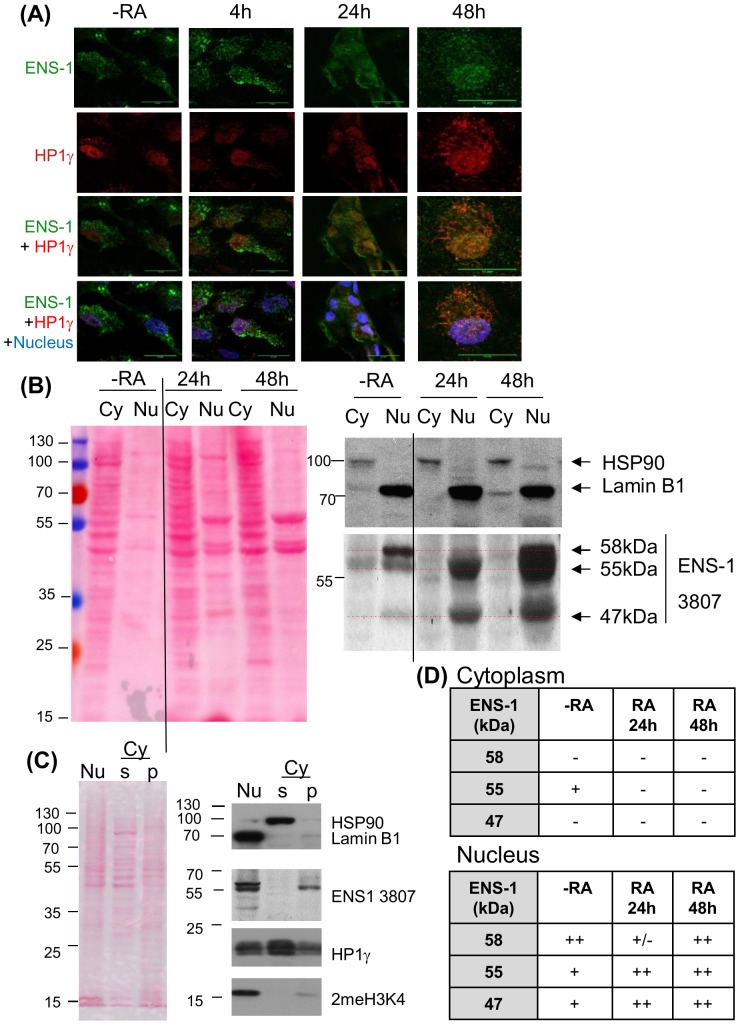
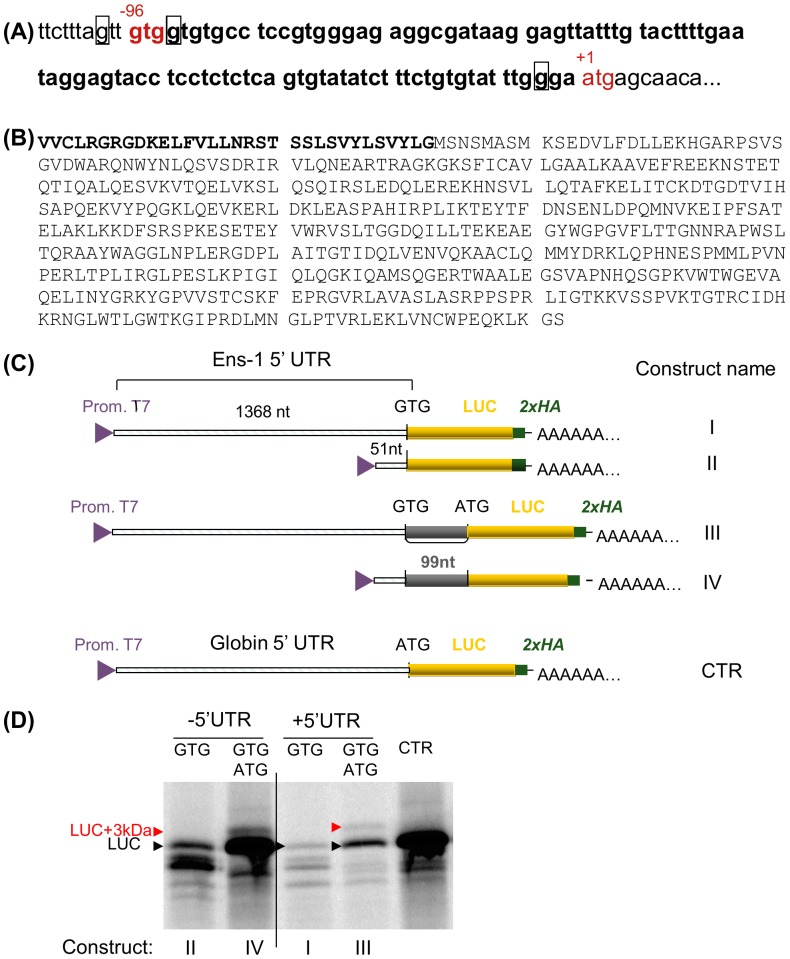
The protein of retroviral origin ENS-1/ERNI plays a major role during neural plate development in chick embryos by controlling the activity of the epigenetic regulator HP1γ, but its function in the earlier developmental stages is still unknown. ENS-1/ERNI promoter activity is down-regulated upon differentiation but the resulting protein expression has never been examined. In this study, we present the results obtained with custom-made antibodies to gain further insights into ENS-1 protein expression in Chicken embryonic stem cells (CES) and during their differentiation. First, we show that ENS-1 controls the activity of HP1γ in CES and we examined the context of its interaction with HP1γ. By combining immunofluorescence and western blot analysis we show that ENS-1 is localized in the cytoplasm and in the nucleus, in agreement with its role on gene's promoter activity. During differentiation, ENS-1 decreases in the cytoplasm but not in the nucleus. More precisely, three distinct forms of the ENS-1 protein co-exist in the nucleus and are differently regulated during differentiation, revealing a new level of control of the protein ENS-1. In silico analysis of the Ens-1 gene copies and the sequence of their corresponding proteins indicate that this pattern is compatible with at least three potential regulation mechanisms, each accounting only partially. The results obtained with the anti-ENS-1 antibodies presented here reveal that the regulation of ENS-1 expression in CES is more complex than expected, providing new tracks to explore the integration of ENS-1 in CES cells regulatory networks.
Conflict of interest statement
Figures

References
-
- Kunarso G, Chia N-Y, Jeyakani J, Hwang C, Lu X, et al. (2010) Transposable elements have rewired the core regulatory network of human embryonic stem cells. Nat Genet 42: 631–634. - PubMed
-
- Lerat E, Birot AM, Samarut J, Mey A (2007) Maintenance in the chicken genome of the retroviral-like cENS gene family specifically expressed in early embryos. J Mol Evol 65: 215–227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
