

Growth and folding of the mammalian cerebral cortex: from molecules to malformations
- PMID: 24646670
- PMCID: PMC4107216
- DOI: 10.1038/nrn3707
Growth and folding of the mammalian cerebral cortex: from molecules to malformations
Abstract
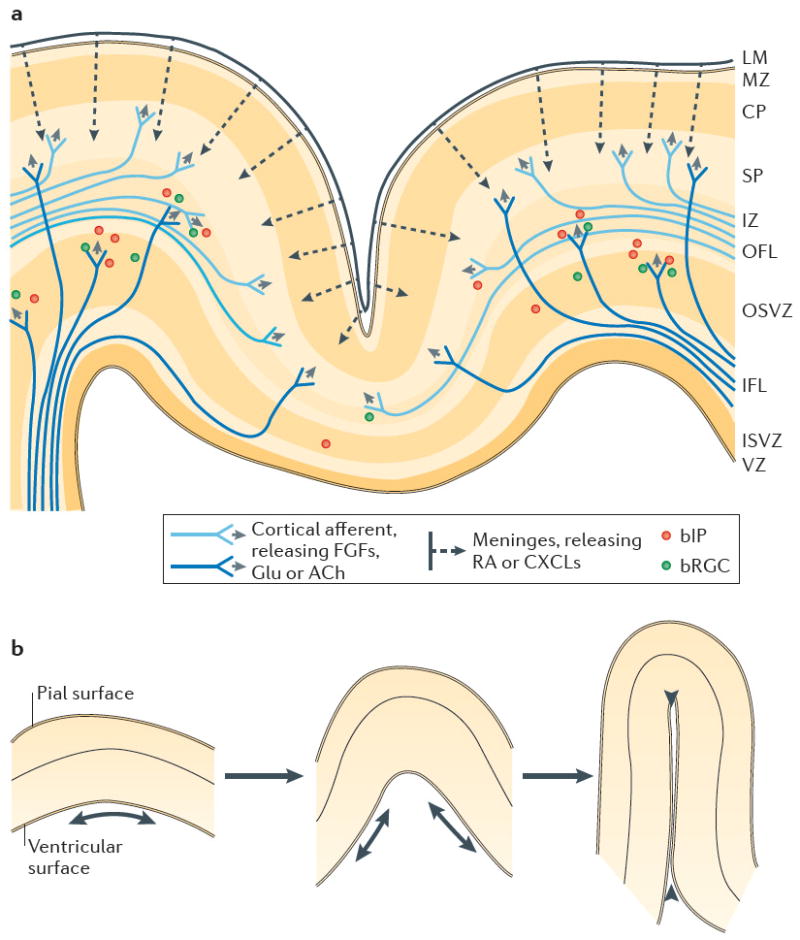
The size and extent of folding of the mammalian cerebral cortex are important factors that influence a species' cognitive abilities and sensorimotor skills. Studies in various animal models and in humans have provided insight into the mechanisms that regulate cortical growth and folding. Both protein-coding genes and microRNAs control cortical size, and recent progress in characterizing basal progenitor cells and the genes that regulate their proliferation has contributed to our understanding of cortical folding. Neurological disorders linked to disruptions in cortical growth and folding have been associated with novel neurogenetic mechanisms and aberrant signalling pathways, and these findings have changed concepts of brain evolution and may lead to new medical treatments for certain disorders.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Striedter G. Principles of Brain Evolution. Sinauer; 2005.
-
- Zilles K, Palomero-Gallagher N, Amunts K. Development of cortical folding during evolution and ontogeny. Trends Neurosci. 2013;36:275–284. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
