

Single-neuron NMDA receptor phenotype influences neuronal rewiring and reintegration following traumatic injury
- PMID: 24647941
- PMCID: PMC3960464
- DOI: 10.1523/JNEUROSCI.4172-13.2014
Single-neuron NMDA receptor phenotype influences neuronal rewiring and reintegration following traumatic injury
Abstract
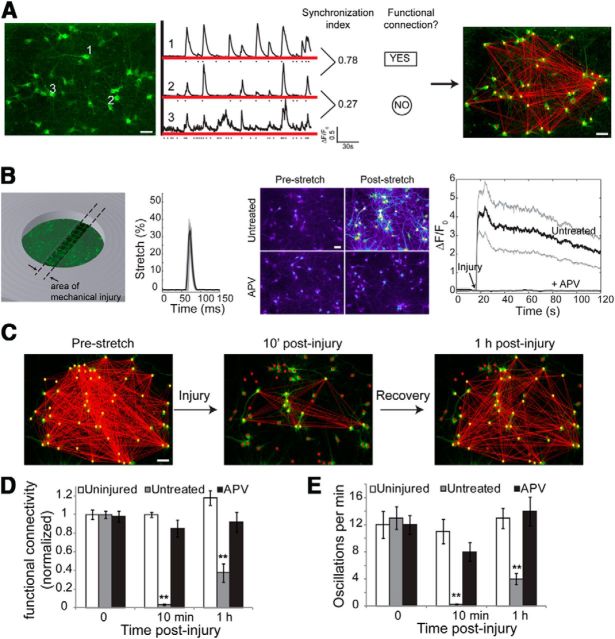
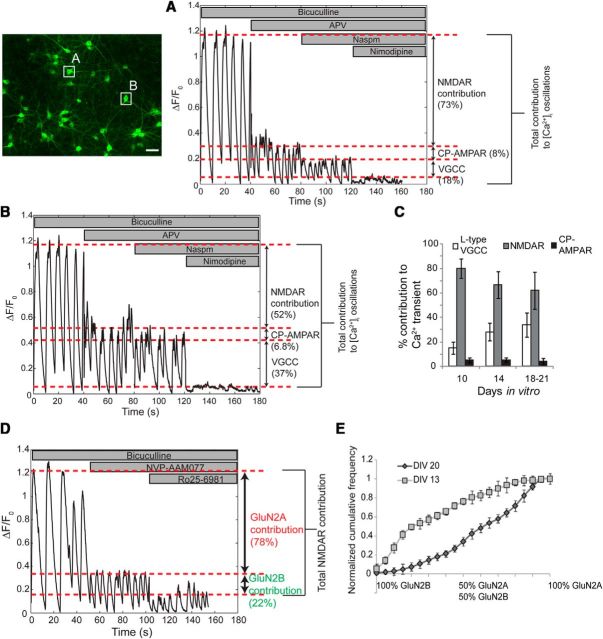
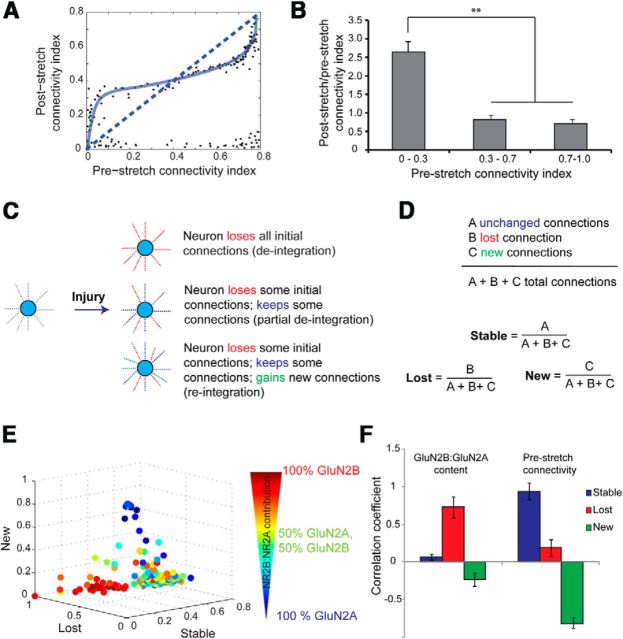
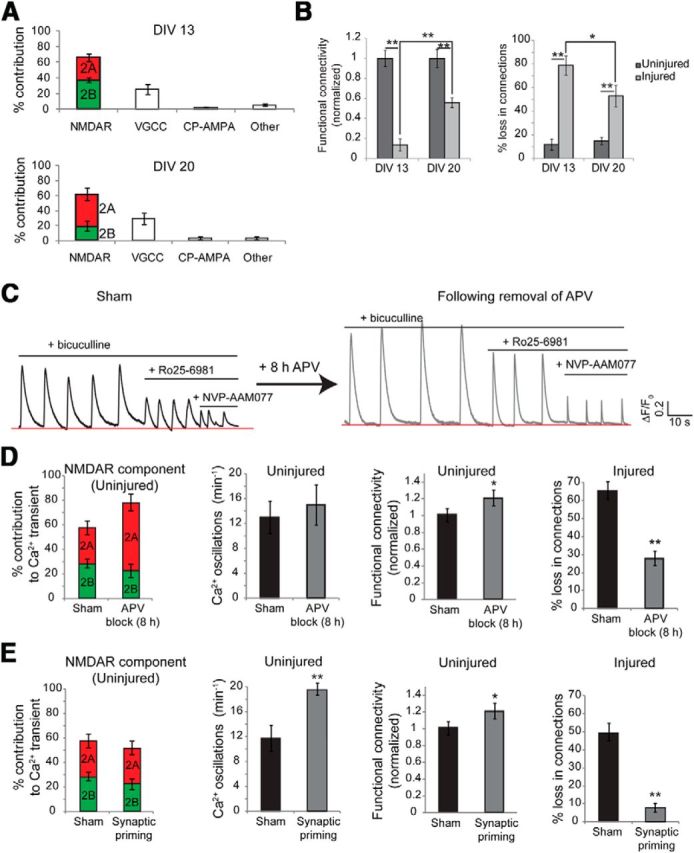
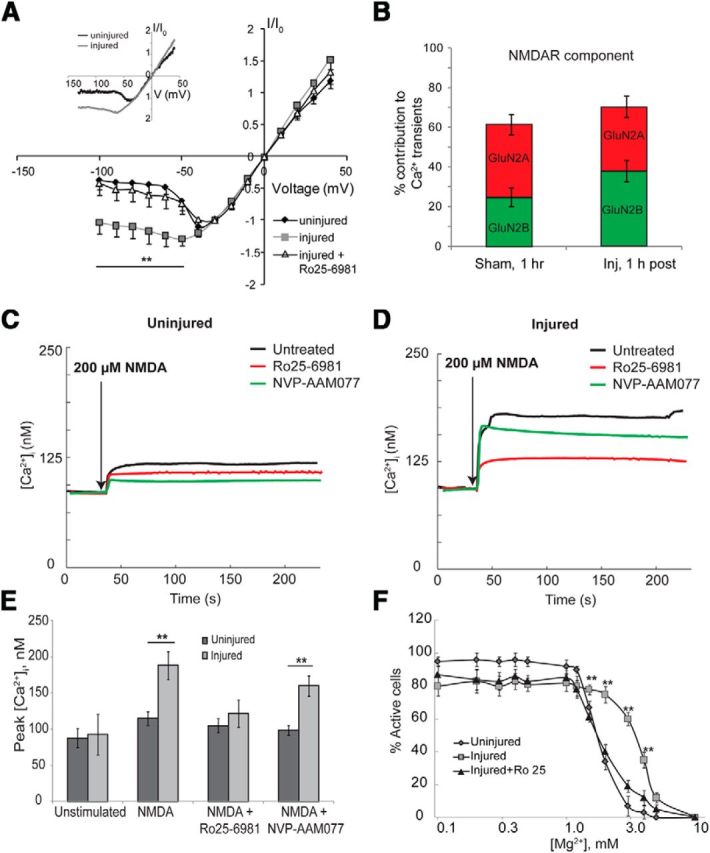
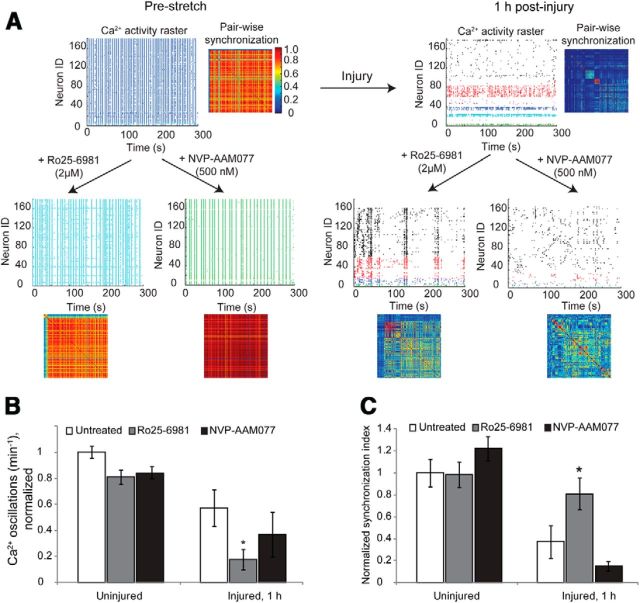
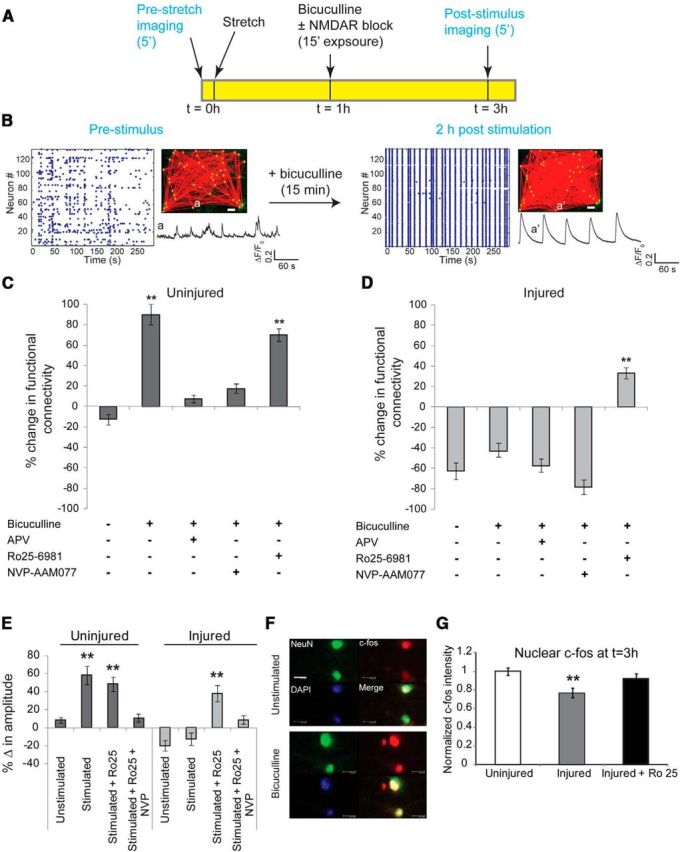
Alterations in the activity of neural circuits are a common consequence of traumatic brain injury (TBI), but the relationship between single-neuron properties and the aggregate network behavior is not well understood. We recently reported that the GluN2B-containing NMDA receptors (NMDARs) are key in mediating mechanical forces during TBI, and that TBI produces a complex change in the functional connectivity of neuronal networks. Here, we evaluated whether cell-to-cell heterogeneity in the connectivity and aggregate contribution of GluN2B receptors to [Ca(2+)]i before injury influenced the functional rewiring, spontaneous activity, and network plasticity following injury using primary rat cortical dissociated neurons. We found that the functional connectivity of a neuron to its neighbors, combined with the relative influx of calcium through distinct NMDAR subtypes, together contributed to the individual neuronal response to trauma. Specifically, individual neurons whose [Ca(2+)]i oscillations were largely due to GluN2B NMDAR activation lost many of their functional targets 1 h following injury. In comparison, neurons with large GluN2A contribution or neurons with high functional connectivity both independently protected against injury-induced loss in connectivity. Mechanistically, we found that traumatic injury resulted in increased uncorrelated network activity, an effect linked to reduction of the voltage-sensitive Mg(2+) block of GluN2B-containing NMDARs. This uncorrelated activation of GluN2B subtypes after injury significantly limited the potential for network remodeling in response to a plasticity stimulus. Together, our data suggest that two single-cell characteristics, the aggregate contribution of NMDAR subtypes and the number of functional connections, influence network structure following traumatic injury.
Keywords: GluN2B; functional connectivity; mechanical injury; network activity; synchrony.
Figures
Similar articles
-
Multiple domains in the C-terminus of NMDA receptor GluN2B subunit contribute to neuronal death following in vitro ischemia.Neurobiol Dis. 2016 May;89:223-34. doi: 10.1016/j.nbd.2015.11.007. Epub 2015 Nov 12. Neurobiol Dis. 2016. PMID: 26581639
-
Amyloid beta peptide 1-42 disturbs intracellular calcium homeostasis through activation of GluN2B-containing N-methyl-d-aspartate receptors in cortical cultures.Cell Calcium. 2012 Feb;51(2):95-106. doi: 10.1016/j.ceca.2011.11.008. Epub 2011 Dec 15. Cell Calcium. 2012. PMID: 22177709
-
Enhanced visual experience rehabilitates the injured brain in Xenopus tadpoles in an NMDAR-dependent manner.J Neurophysiol. 2019 Jan 1;121(1):306-320. doi: 10.1152/jn.00664.2018. Epub 2018 Dec 5. J Neurophysiol. 2019. PMID: 30517041 Free PMC article.
-
Role of nonsynaptic GluN2B-containing NMDA receptors in excitotoxicity: evidence that fluoxetine selectively inhibits these receptors and may have neuroprotective effects.Brain Res Bull. 2013 Apr;93:32-8. doi: 10.1016/j.brainresbull.2012.10.005. Epub 2012 Oct 23. Brain Res Bull. 2013. PMID: 23089362 Review.
-
NMDA receptor hypofunction, parvalbumin-positive neurons, and cortical gamma oscillations in schizophrenia.Schizophr Bull. 2012 Sep;38(5):950-7. doi: 10.1093/schbul/sbs010. Epub 2012 Feb 21. Schizophr Bull. 2012. PMID: 22355184 Free PMC article. Review.
Cited by
-
Revisiting Traumatic Brain Injury: From Molecular Mechanisms to Therapeutic Interventions.Biomedicines. 2020 Sep 29;8(10):389. doi: 10.3390/biomedicines8100389. Biomedicines. 2020. PMID: 33003373 Free PMC article. Review.
-
Mild traumatic brain injury as a pathological process.Heliyon. 2023 Jul 17;9(7):e18342. doi: 10.1016/j.heliyon.2023.e18342. eCollection 2023 Jul. Heliyon. 2023. PMID: 37519712 Free PMC article. Review.
-
High-Frequency Head Impact Disrupts Hippocampal Neural Ensemble Dynamics.Front Cell Neurosci. 2022 Jan 18;15:763423. doi: 10.3389/fncel.2021.763423. eCollection 2021. Front Cell Neurosci. 2022. PMID: 35115908 Free PMC article.
-
Proposed mechanisms of tau: relationships to traumatic brain injury, Alzheimer's disease, and epilepsy.Front Neurol. 2024 Jan 5;14:1287545. doi: 10.3389/fneur.2023.1287545. eCollection 2023. Front Neurol. 2024. PMID: 38249745 Free PMC article.
-
Comparative Efficacy and Safety of 11 Drugs as Therapies for Adults With Neuropathic Pain After Spinal Cord Injury: A Bayesian Network Analysis Based on 20 Randomized Controlled Trials.Front Neurol. 2022 Mar 21;13:818522. doi: 10.3389/fneur.2022.818522. eCollection 2022. Front Neurol. 2022. PMID: 35386408 Free PMC article.
References
-
- Akerboom J, Chen TW, Wardill TJ, Tian L, Marvin JS, Mutlu S, Calderón NC, Esposti F, Borghuis BG, Sun XR, Gordus A, Orger MB, Portugues R, Engert F, Macklin JJ, Filosa A, Aggarwal A, Kerr RA, Takagi R, Kracun S, et al. Optimization of a GCaMP calcium indicator for neural activity imaging. J Neurosci. 2012;32:13819–13840. doi: 10.1523/JNEUROSCI.2601-12.2012. - DOI - PMC - PubMed
-
- Arnold FJ, Hofmann F, Bengtson CP, Wittmann M, Vanhoutte P, Bading H. Microelectrode array recordings of cultured hippocampal networks reveal a simple model for transcription and protein synthesis-dependent plasticity. J Physiol. 2005;564:3–19. doi: 10.1113/jphysiol.2004.077446. - DOI - PMC - PubMed
-
- Auberson YP, Allgeier H, Bischoff S, Lingenhoehl K, Moretti R, Schmutz M. 5-Phosphonomethylquinoxalinediones as competitive NMDA receptor antagonists with a preference for the human 1A/2A, rather than 1A/2B receptor composition. Bioorg Med Chem Lett. 2002;12:1099–1102. doi: 10.1016/S0960-894X(02)00074-4. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous