

Organization and execution of the epithelial polarity programme
- PMID: 24651541
- PMCID: PMC4211427
- DOI: 10.1038/nrm3775
Organization and execution of the epithelial polarity programme
Abstract
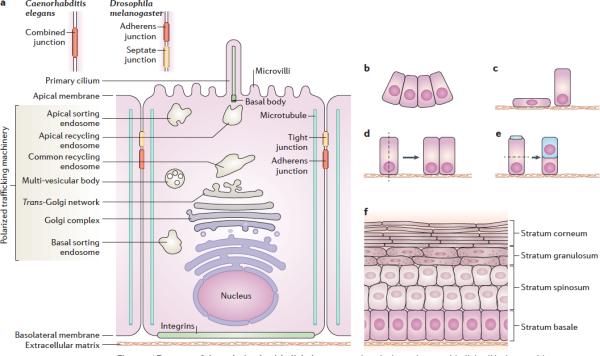
Epithelial cells require apical-basal plasma membrane polarity to carry out crucial vectorial transport functions and cytoplasmic polarity to generate different cell progenies for tissue morphogenesis. The establishment and maintenance of a polarized epithelial cell with apical, basolateral and ciliary surface domains is guided by an epithelial polarity programme (EPP) that is controlled by a network of protein and lipid regulators. The EPP is organized in response to extracellular cues and is executed through the establishment of an apical-basal axis, intercellular junctions, epithelial-specific cytoskeletal rearrangements and a polarized trafficking machinery. Recent studies have provided insight into the interactions of the EPP with the polarized trafficking machinery and how these regulate epithelial polarization and depolarization.
Figures




References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
