

Metabolic requirements for the maintenance of self-renewing stem cells
- PMID: 24651542
- PMCID: PMC4095859
- DOI: 10.1038/nrm3772
Metabolic requirements for the maintenance of self-renewing stem cells
Abstract
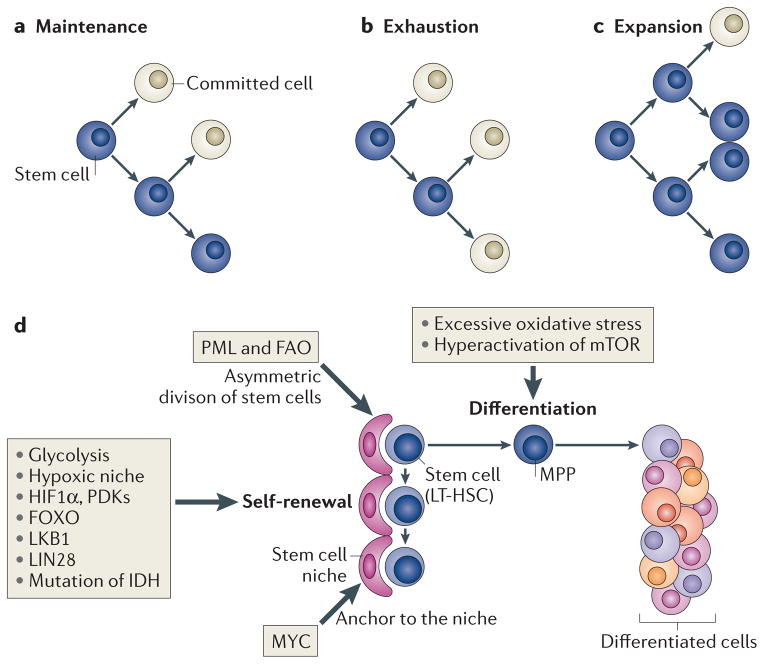
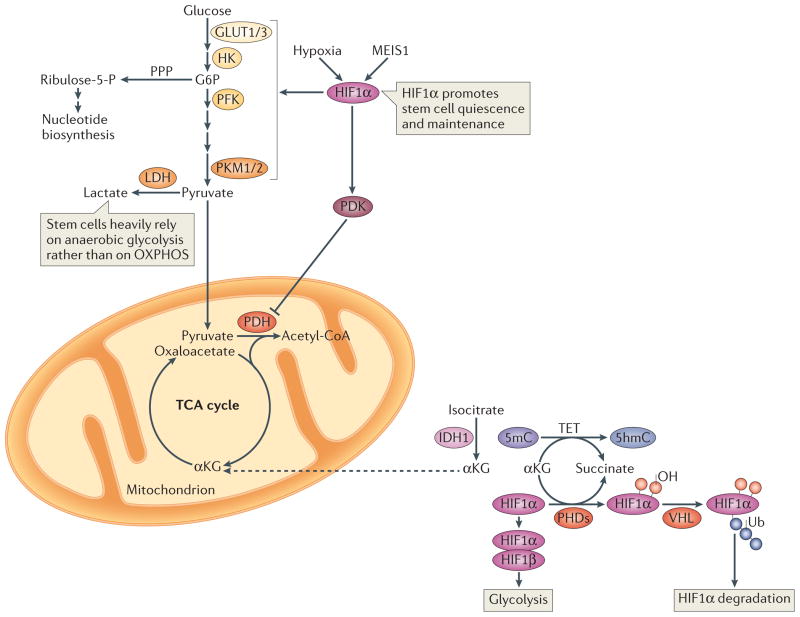
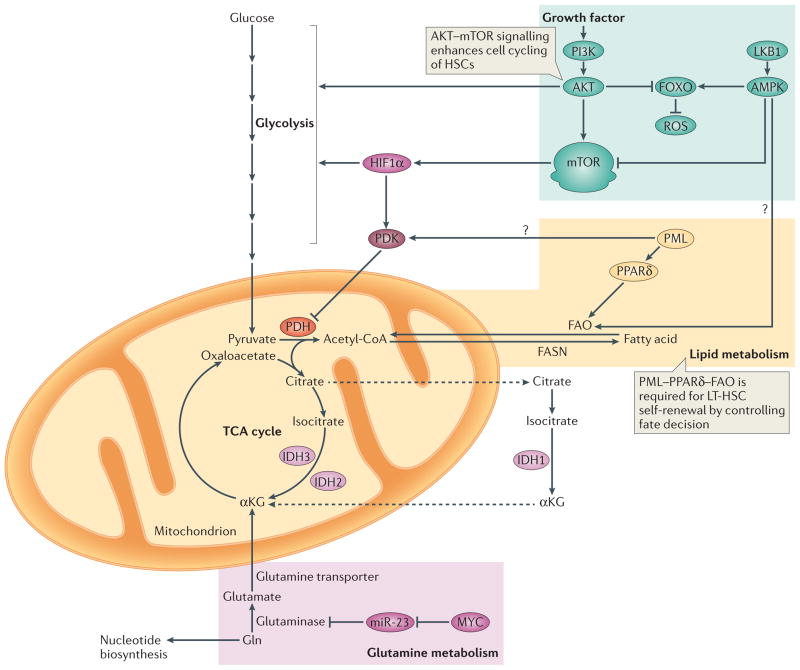
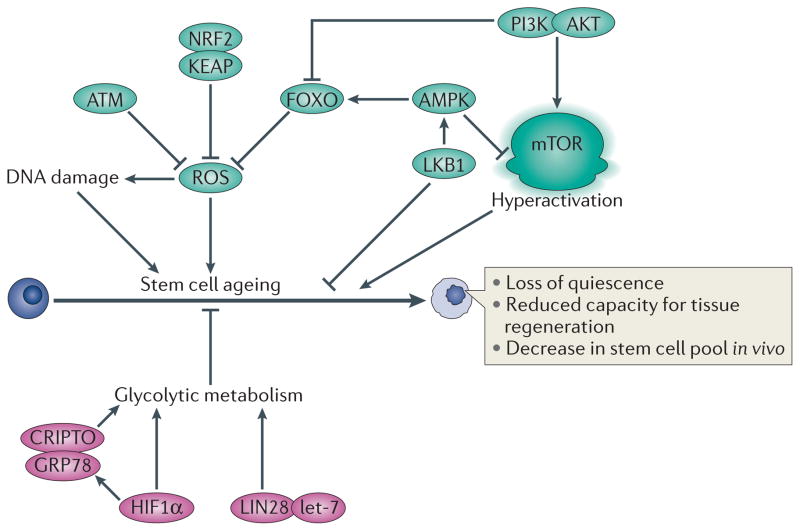
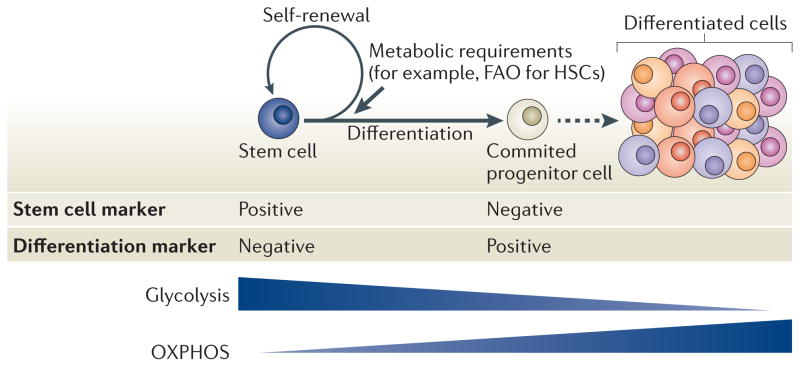
A distinctive feature of stem cells is their capacity to self-renew to maintain pluripotency. Studies of genetically-engineered mouse models and recent advances in metabolomic analysis, particularly in haematopoietic stem cells, have deepened our understanding of the contribution made by metabolic cues to the regulation of stem cell self-renewal. Many types of stem cells heavily rely on anaerobic glycolysis, and stem cell function is also regulated by bioenergetic signalling, the AKT-mTOR pathway, Gln metabolism and fatty acid metabolism. As maintenance of a stem cell pool requires a finely-tuned balance between self-renewal and differentiation, investigations into the molecular mechanisms and metabolic pathways underlying these decisions hold great therapeutic promise.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Weissman IL, Anderson DJ, Gage F. Stem and progenitor cells: origins, phenotypes, lineage commitments, and transdifferentiations. Annu Rev Cell Dev Biol. 2001;17:387–403. - PubMed
-
- Zon LI. Intrinsic and extrinsic control of haematopoietic stem-cell self-renewal. Nature. 2008;453:306–313. - PubMed
-
- He S, Nakada D, Morrison SJ. Mechanisms of stem cell self-renewal. Annu Rev Cell Dev Biol. 2009;25:377–406. - PubMed
-
- van der Lugt NM, et al. Posterior transformation, neurological abnormalities, and severe hematopoietic defects in mice with a targeted deletion of the bmi-1 proto-oncogene. Genes Dev. 1994;8:757–769. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
