

Activation of 2' 5'-oligoadenylate synthetase by stem loops at the 5'-end of the West Nile virus genome
- PMID: 24651762
- PMCID: PMC3961380
- DOI: 10.1371/journal.pone.0092545
Activation of 2' 5'-oligoadenylate synthetase by stem loops at the 5'-end of the West Nile virus genome
Abstract
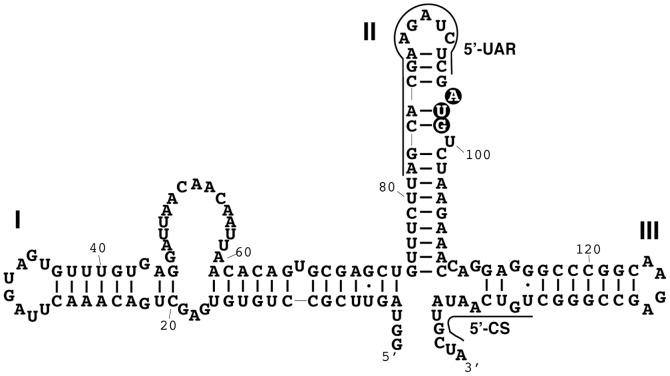
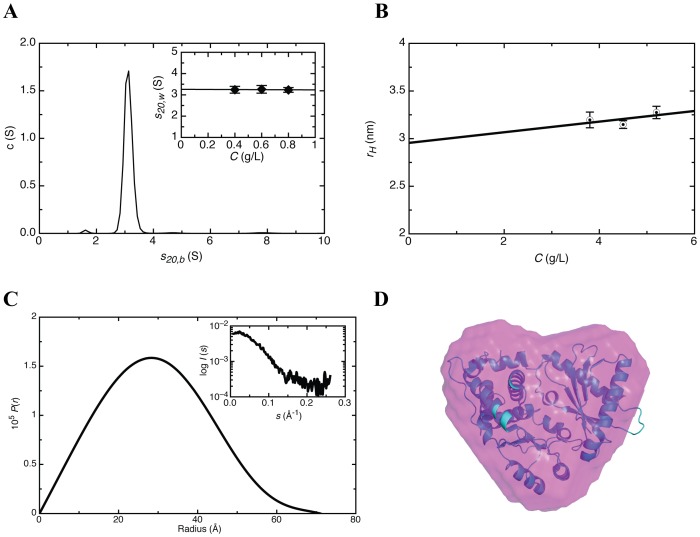
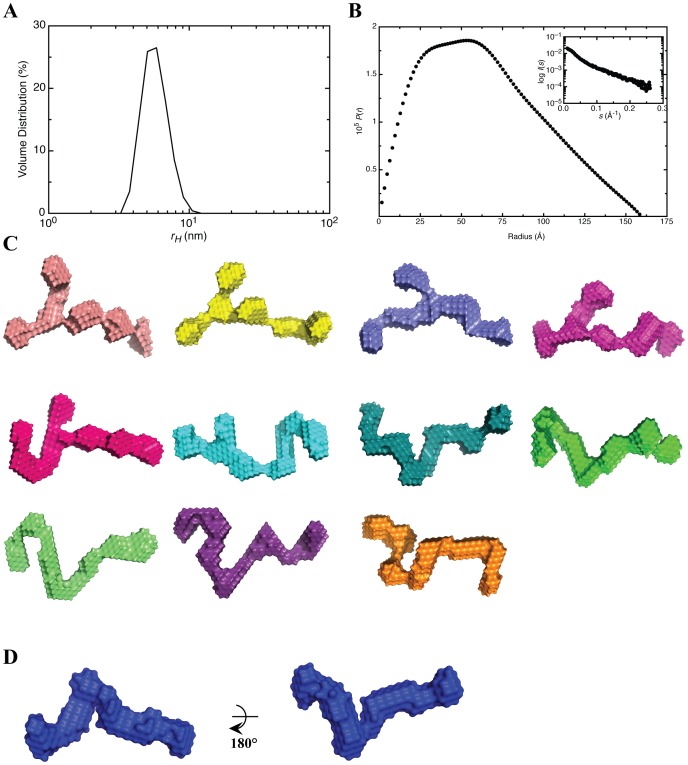
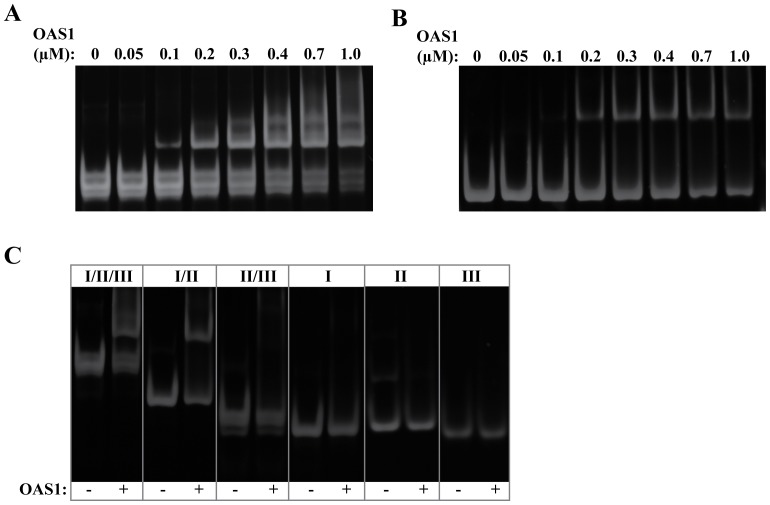
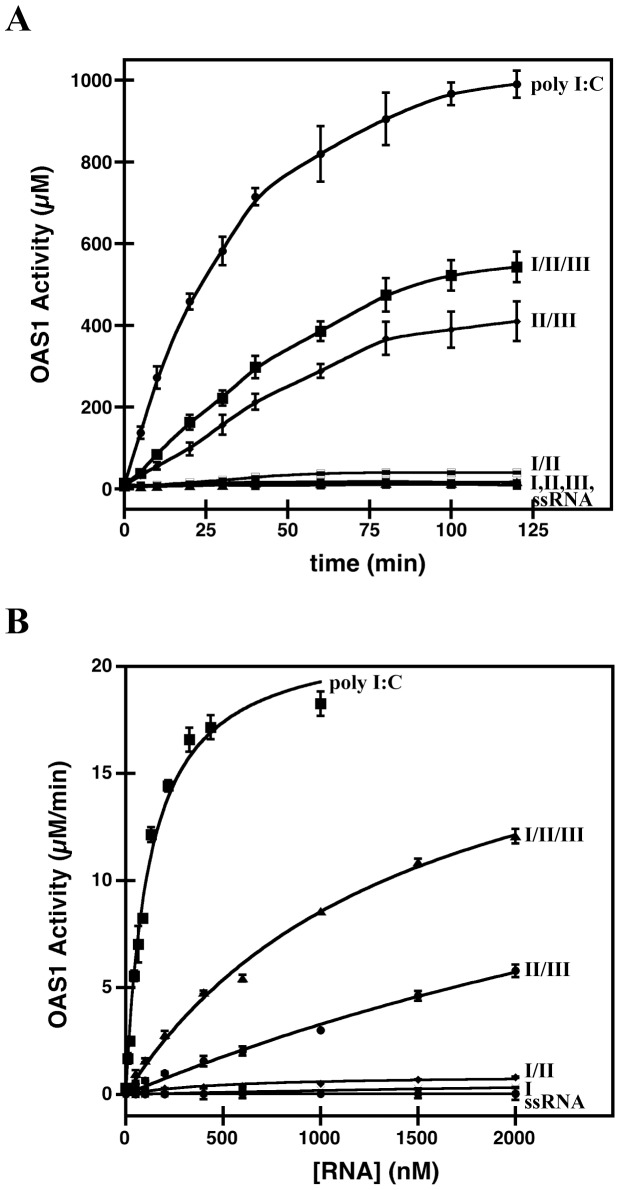
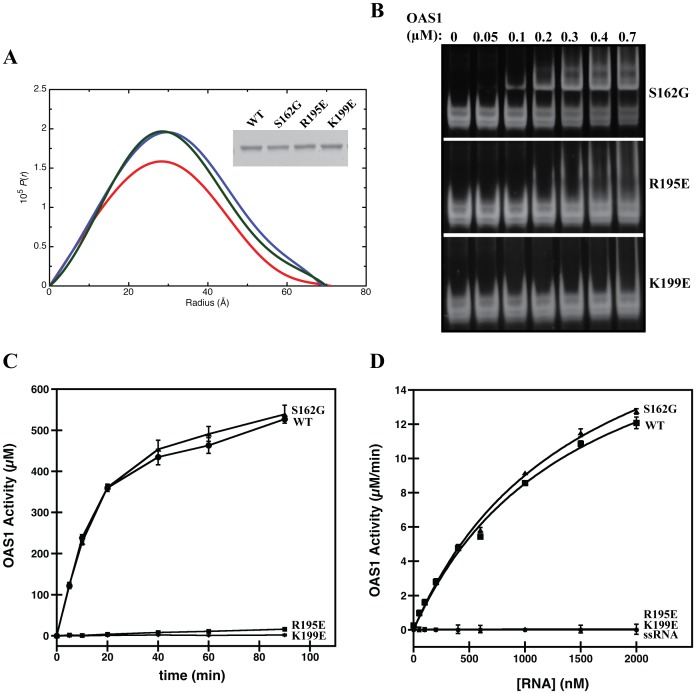
West Nile virus (WNV) has a positive sense RNA genome with conserved structural elements in the 5' and 3' -untranslated regions required for polyprotein production. Antiviral immunity to WNV is partially mediated through the production of a cluster of proteins known as the interferon stimulated genes (ISGs). The 2' 5'-oligoadenylate synthetases (OAS) are key ISGs that help to amplify the innate immune response. Upon interaction with viral double stranded RNA, OAS enzymes become activated and enable the host cell to restrict viral propagation. Studies have linked mutations in the OAS1 gene to increased susceptibility to WNV infection, highlighting the importance of OAS1 enzyme. Here we report that the region at the 5'-end of the WNV genome comprising both the 5'-UTR and initial coding region is capable of OAS1 activation in vitro. This region contains three RNA stem loops (SLI, SLII, and SLIII), whose relative contribution to OAS1 binding affinity and activation were investigated using electrophoretic mobility shift assays and enzyme kinetics experiments. Stem loop I, comprising nucleotides 1-73, is dispensable for maximum OAS1 activation, as a construct containing only SLII and SLIII was capable of enzymatic activation. Mutations to the RNA binding site of OAS1 confirmed the specificity of the interaction. The purity, monodispersity and homogeneity of the 5'-end (SLI/II/III) and OAS1 were evaluated using dynamic light scattering and analytical ultra-centrifugation. Solution conformations of both the 5'-end RNA of WNV and OAS1 were then elucidated using small-angle x-ray scattering. In the context of purified components in vitro, these data demonstrate the recognition of conserved secondary structural elements of the WNV genome by a member of the interferon-mediated innate immune response.
Conflict of interest statement
Figures
References
-
- Gubler DJ (2007) The continuing spread of West Nile virus in the western hemisphere. Clin Infect Dis 45: 1039–1046. - PubMed
-
- Asnis DS, Conetta R, Teixeira AA, Waldman G, Sampson BA (2000) The West Nile virus outbreak of 1999 in New York: the Flushing Hospital experience. (vol 30, pg 413, 2000). Clinical Infectious Diseases 30: 841–841. - PubMed
-
- Savage HM, Ceianu C, Nicolescu G, Karabatsos N, Lanciotti R, et al. (1999) Entomologic and avian investigations of an epidemic of West Nile fever in Romania in 1996, with serologic and molecular characterization of a virus isolate from mosquitoes. Am J Trop Med Hyg 61: 600–611. - PubMed
-
- Lanciotti RS, Ebel GD, Deubel V, Kerst AJ, Murri S, et al. (2002) Complete genome sequences and phylogenetic analysis of West Nile virus strains isolated from the United States, Europe, and the Middle East. Virology 298: 96–105. - PubMed
-
- Calisher CH (2000) West Nile virus in the New World: appearance, persistence, and adaptation to a new econiche—an opportunity taken. Viral Immunol 13: 411–414. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
