

Co-factor insertion and disulfide bond requirements for twin-arginine translocase-dependent export of the Bacillus subtilis Rieske protein QcrA
- PMID: 24652282
- PMCID: PMC4036324
- DOI: 10.1074/jbc.M113.529677
Co-factor insertion and disulfide bond requirements for twin-arginine translocase-dependent export of the Bacillus subtilis Rieske protein QcrA
Abstract
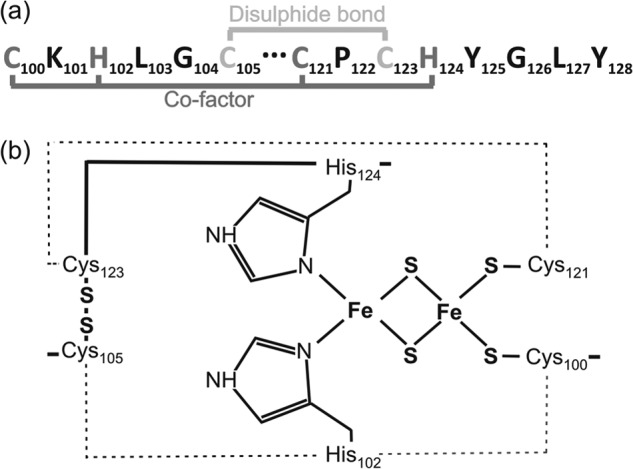
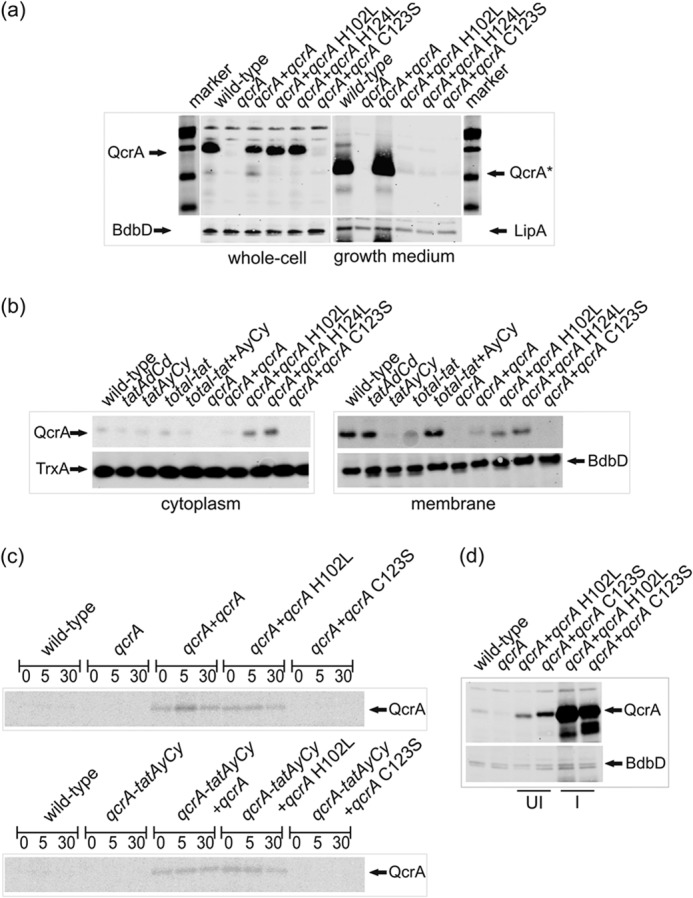
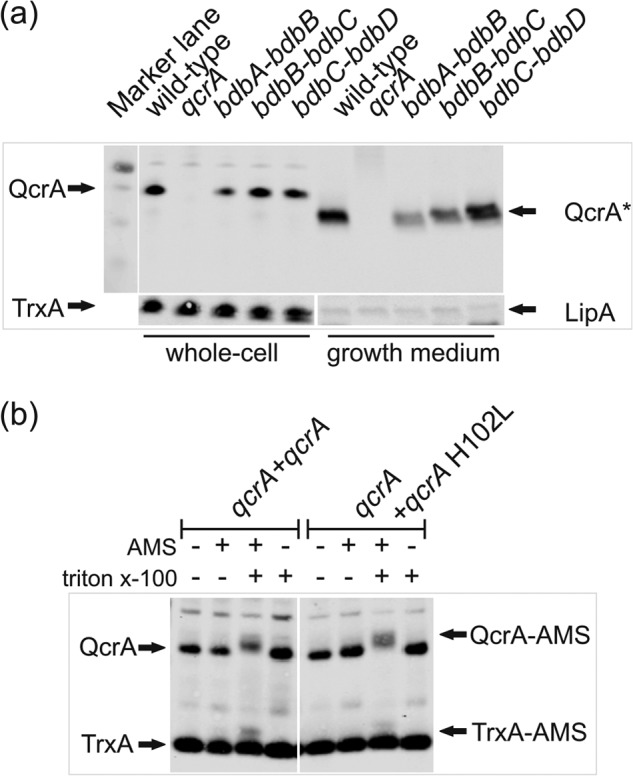
The twin-arginine translocation (Tat) pathway can transport folded and co-factor-containing cargo proteins over bacterial cytoplasmic membranes. Functional Tat machinery components, a folded state of the cargo protein and correct co-factor insertion in the cargo protein are generally considered as prerequisites for successful translocation. The present studies were aimed at a dissection of these requirements with regard to the Rieske iron-sulfur protein QcrA of Bacillus subtilis. Notably, QcrA is a component of the cytochrome bc1 complex, which is conserved from bacteria to man. Single amino acid substitutions were introduced into the Rieske domain of QcrA to prevent either co-factor binding or disulfide bond formation. Both types of mutations precluded QcrA translocation. Importantly, a proofreading hierarchy was uncovered, where a QcrA mutant defective in disulfide bonding was quickly degraded, whereas mutant QcrA proteins defective in co-factor binding accumulated in the cytoplasm and membrane. Altogether, these are the first studies on Tat-dependent protein translocation where both oxidative folding and co-factor attachment have been addressed in a single native molecule.
Keywords: Bacillus; Cytochromes; Disulfide; Iron-Sulfur Protein; Protein Translocation; QcrA; Rieske Protein; TatAy; Twin-arginine Translocation.
Figures
Similar articles
-
A Tat ménage à trois--The role of Bacillus subtilis TatAc in twin-arginine protein translocation.Biochim Biophys Acta. 2015 Oct;1853(10 Pt A):2745-53. doi: 10.1016/j.bbamcr.2015.07.022. Epub 2015 Aug 1. Biochim Biophys Acta. 2015. PMID: 26239117
-
Role of the twin arginine protein transport pathway in the assembly of the Streptomyces coelicolor cytochrome bc1 complex.J Bacteriol. 2014 Jan;196(1):50-9. doi: 10.1128/JB.00776-13. Epub 2013 Oct 18. J Bacteriol. 2014. PMID: 24142258 Free PMC article.
-
Double trouble: Bacillus depends on a functional Tat machinery to avoid severe oxidative stress and starvation upon entry into a NaCl-depleted environment.Biochim Biophys Acta Mol Cell Res. 2021 Feb;1868(2):118914. doi: 10.1016/j.bbamcr.2020.118914. Epub 2020 Nov 25. Biochim Biophys Acta Mol Cell Res. 2021. PMID: 33245978
-
The Tat system of Gram-positive bacteria.Biochim Biophys Acta. 2014 Aug;1843(8):1698-706. doi: 10.1016/j.bbamcr.2013.10.008. Epub 2013 Oct 16. Biochim Biophys Acta. 2014. PMID: 24140208 Review.
-
Twin-Arginine Protein Translocation.Curr Top Microbiol Immunol. 2017;404:69-94. doi: 10.1007/82_2016_7. Curr Top Microbiol Immunol. 2017. PMID: 27121927 Review.
Cited by
-
Targeting of proteins to the twin-arginine translocation pathway.Mol Microbiol. 2020 May;113(5):861-871. doi: 10.1111/mmi.14461. Epub 2020 Feb 20. Mol Microbiol. 2020. PMID: 31971282 Free PMC article. Review.
-
The Twin-Arginine Pathway for Protein Secretion.EcoSal Plus. 2019 Jun;8(2):10.1128/ecosalplus.ESP-0040-2018. doi: 10.1128/ecosalplus.ESP-0040-2018. EcoSal Plus. 2019. PMID: 31215506 Free PMC article. Review.
-
A TatABC-type Tat translocase is required for unimpaired aerobic growth of Corynebacterium glutamicum ATCC13032.PLoS One. 2015 Apr 2;10(4):e0123413. doi: 10.1371/journal.pone.0123413. eCollection 2015. PLoS One. 2015. PMID: 25837592 Free PMC article.
-
Transport of Folded Proteins by the Tat System.Protein J. 2019 Aug;38(4):377-388. doi: 10.1007/s10930-019-09859-y. Protein J. 2019. PMID: 31401776 Free PMC article. Review.
References
-
- Maurer C., Panahandeh S., Moser M., Müller M. (2009) Impairment of twin-arginine-dependent export by seemingly small alterations of substrate conformation. FEBS Lett. 583, 2849–2853 - PubMed
-
- Oresnik I. J., Ladner C. L., Turner R. J. (2001) Identification of a twin-arginine leader-binding protein. Mol. Microbiol. 40, 323–331 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
