

Dynamic self-assembly of motile bacteria in liquid crystals
- PMID: 24652584
- PMCID: PMC3966026
- DOI: 10.1039/c3sm52423j
Dynamic self-assembly of motile bacteria in liquid crystals
Abstract
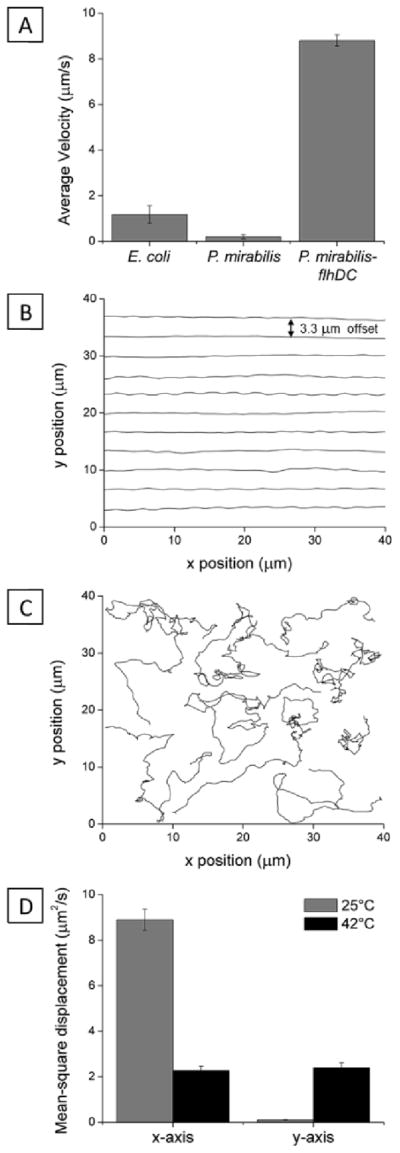
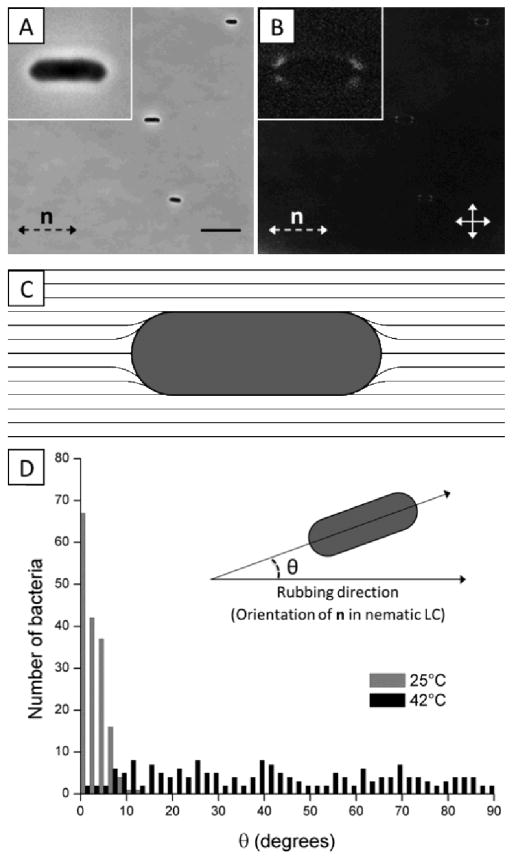
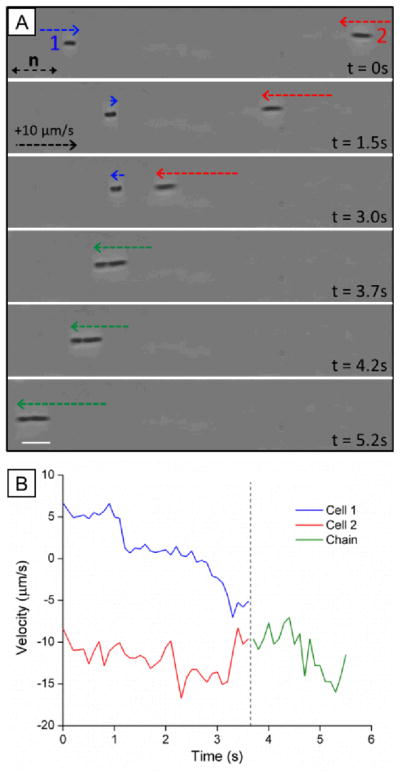
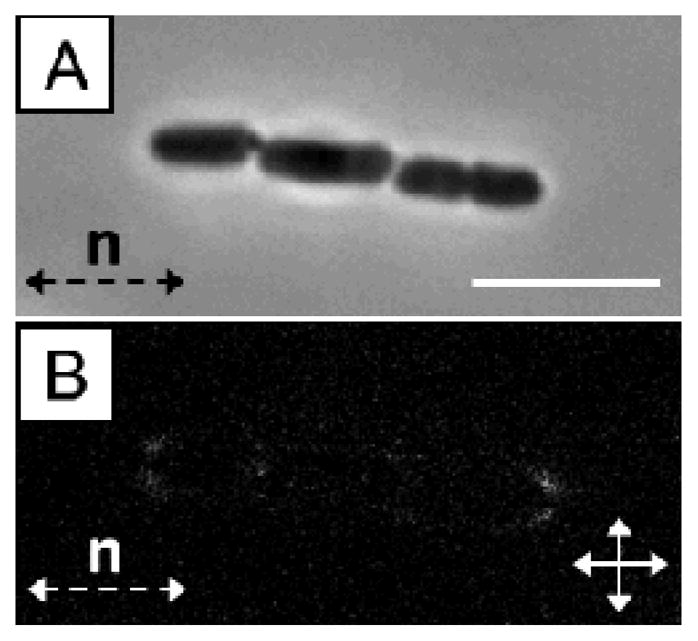
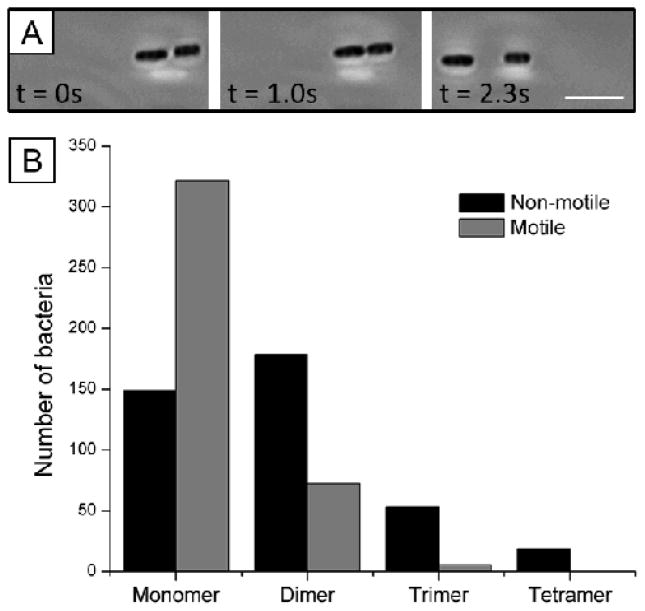
This paper reports an investigation of dynamical behaviors of motile rod-shaped bacteria within anisotropic viscoelastic environments defined by lyotropic liquid crystals (LCs). In contrast to passive microparticles (including non-motile bacteria) that associate irreversibly in LCs via elasticity-mediated forces, we report that motile Proteus mirabilis bacteria form dynamic and reversible multi-cellular assemblies when dispersed in a lyotropic LC. By measuring the velocity of the bacteria through the LC (8.8 ± 0.2 μm s(-1)) and by characterizing the ordering of the LC about the rod-shaped bacteria (tangential anchoring), we conclude that the reversibility of the inter-bacterial interaction emerges from the interplay of forces generated by the flagella of the bacteria and the elasticity of the LC, both of which are comparable in magnitude (tens of pN) for motile Proteus mirabilis cells. We also measured the dissociation process, which occurs in a direction determined by the LC, to bias the size distribution of multi-cellular bacterial complexes in a population of motile Proteus mirabilis relative to a population of non-motile cells. Overall, these observations and others reported in this paper provide insight into the fundamental dynamic behaviors of bacteria in complex anisotropic environments and suggest that motile bacteria in LCs are an exciting model system for exploration of principles for the design of active materials.
Conflict of interest statement
The authors declare the following competing financial interest(s): NLA declares a significant financial interest in Platypus Technologies LLC, a for-profit company that has developed LC-based technologies for molecular analysis.
Figures
References
-
- Shaw T, Winston M, Rupp CJ, Klapper I, Stoodley P. Phys Rev Lett. 2004;93:098102. - PubMed
-
- Madoff LC, Thaler SJ, Maguire JH. In: Harrison’s Principles of Internal Medicine. 16. Braunwald E, Fauci AS, Kasper DL, Hauser SL, Longo DL, Jameson JL, editors. McGraw-Hill; New York: 2005. pp. 2050–2055.
-
- Flemming HC, Wingender J. Nat Rev Microbiol. 2010;8:623–633. - PubMed
-
- Rey AD. Soft Matter. 2010;6:3402–3429.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources