

Translational profiles of medullary myofibroblasts during kidney fibrosis
- PMID: 24652793
- PMCID: PMC4147988
- DOI: 10.1681/ASN.2013101143
Translational profiles of medullary myofibroblasts during kidney fibrosis
Abstract
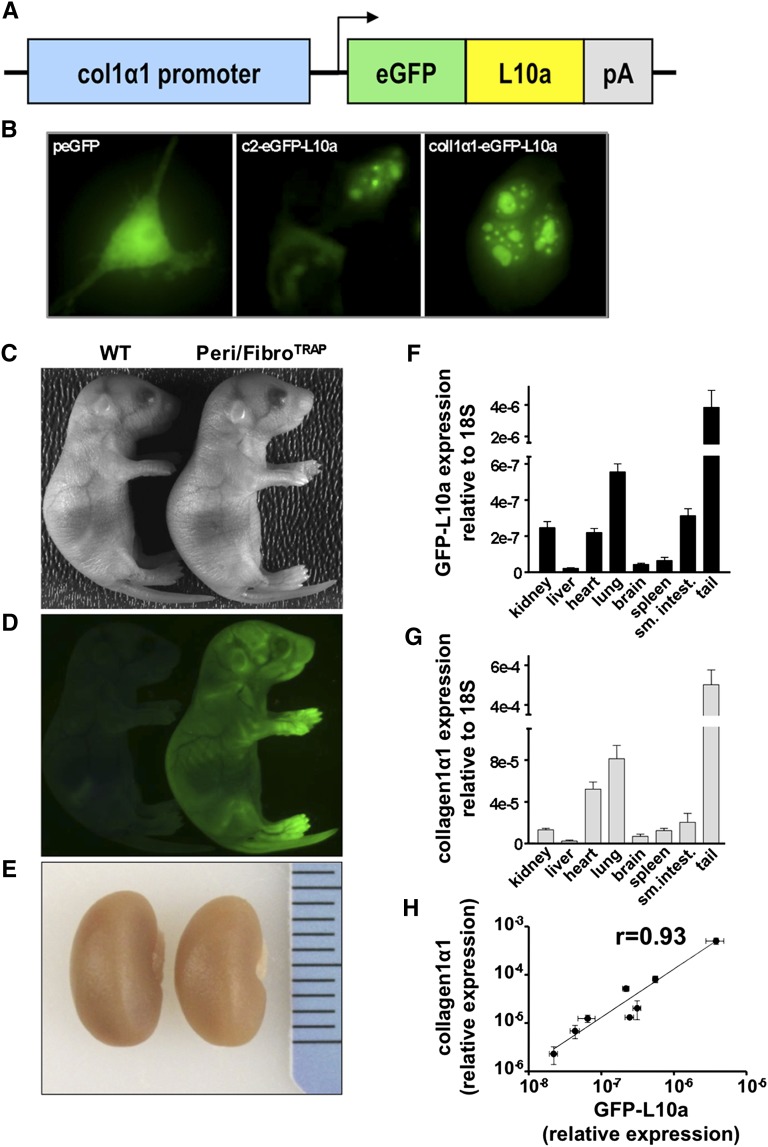
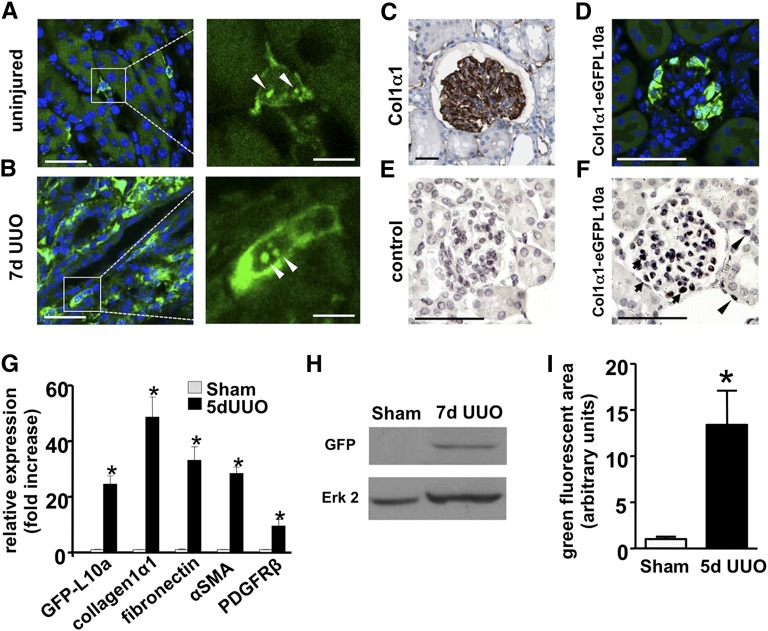
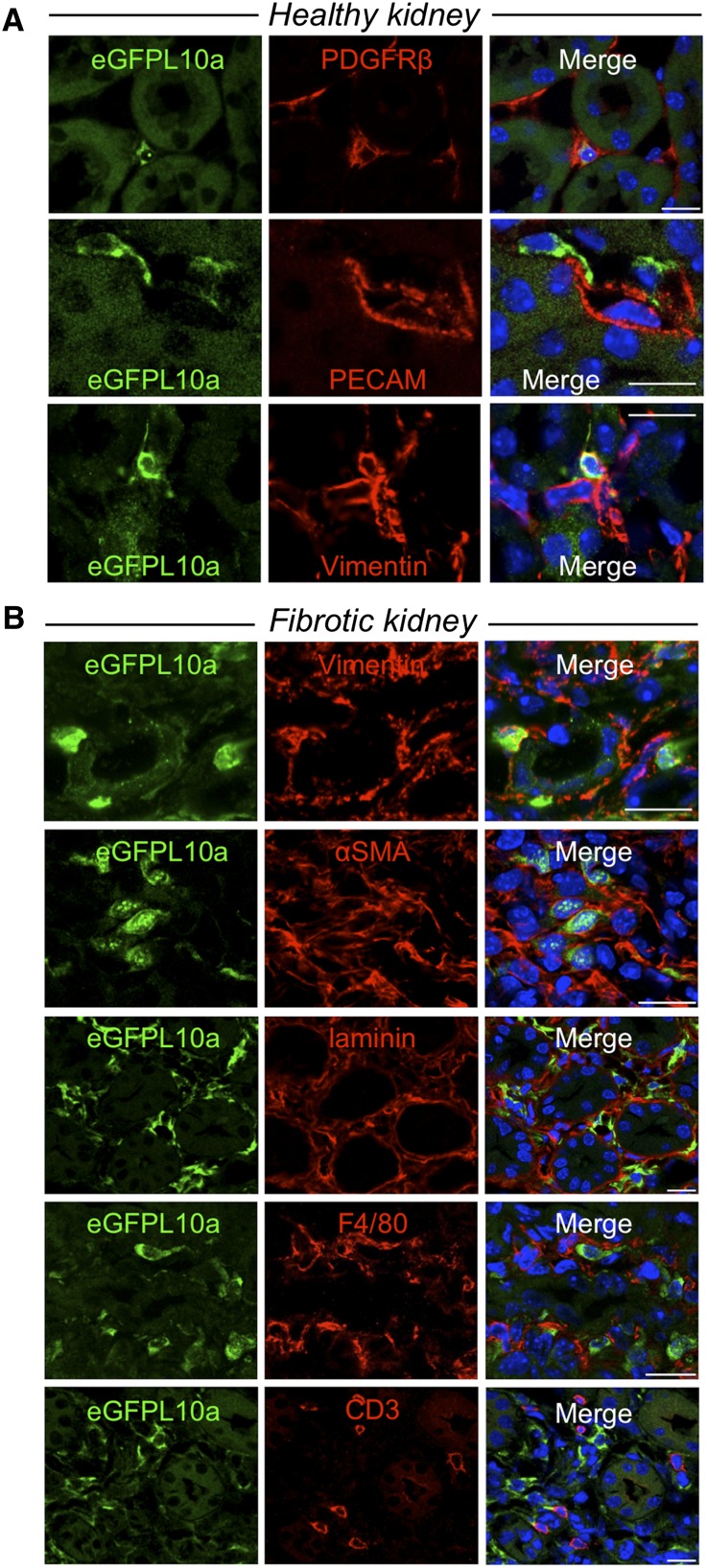
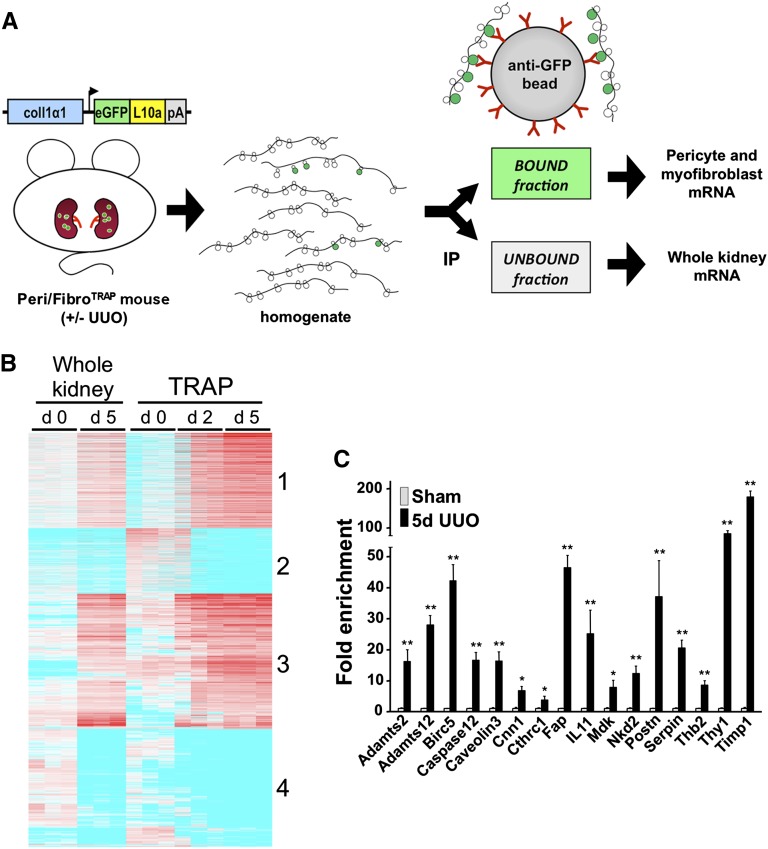
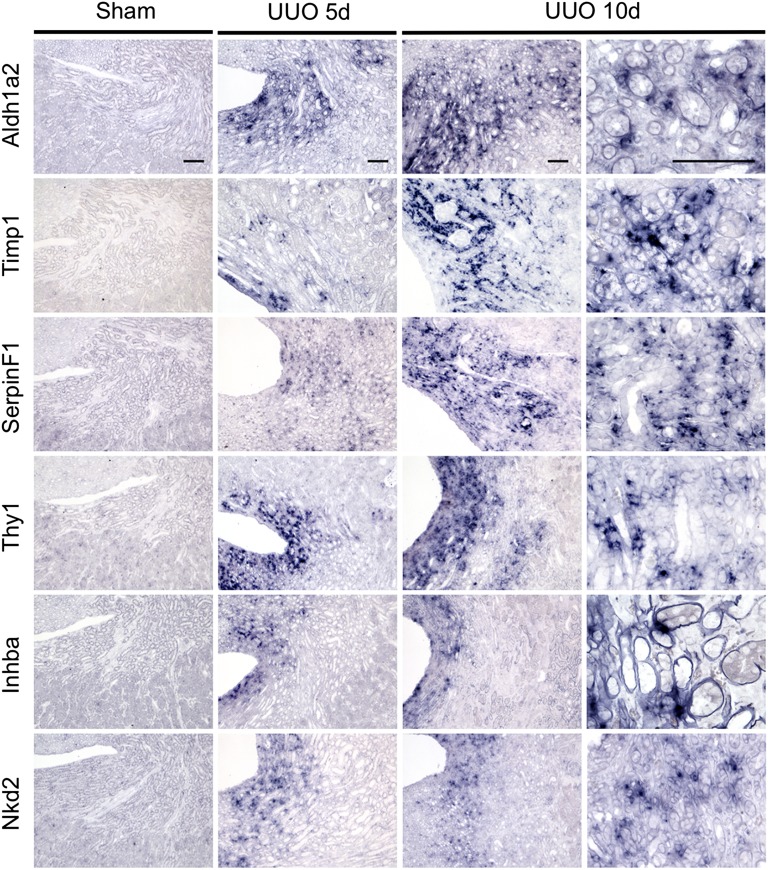
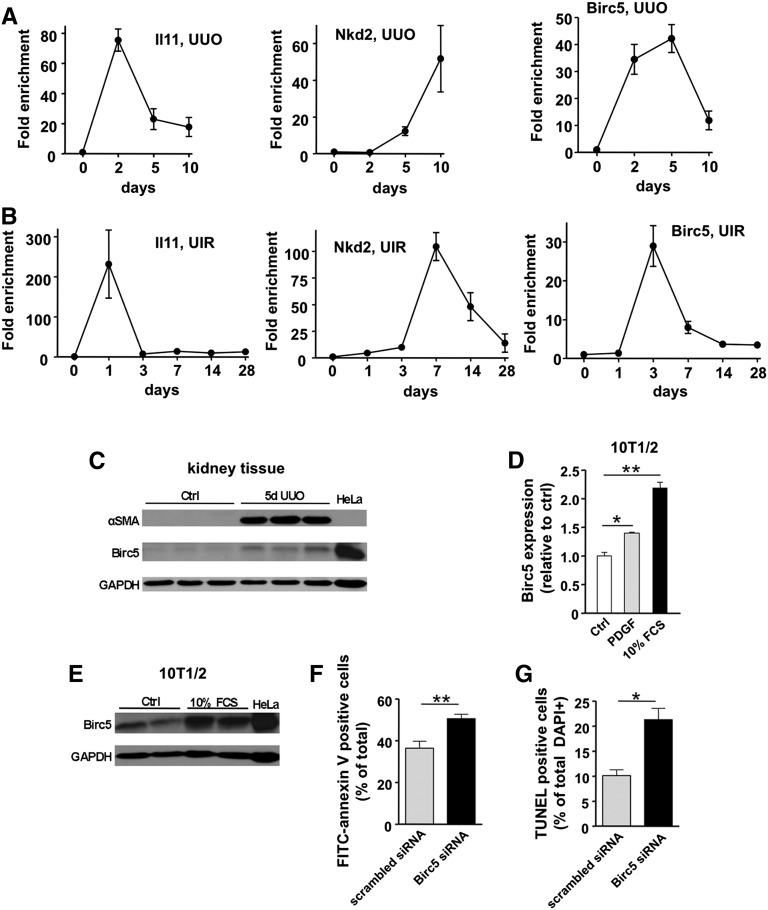
Myofibroblasts secrete matrix during chronic injury, and their ablation ameliorates fibrosis. Development of new biomarkers and therapies for CKD will be aided by a detailed analysis of myofibroblast gene expression during the early stages of fibrosis. However, dissociating myofibroblasts from fibrotic kidney is challenging. We therefore adapted translational ribosome affinity purification (TRAP) to isolate and profile mRNA from myofibroblasts and their precursors during kidney fibrosis. We generated and characterized a transgenic mouse expressing an enhanced green fluorescent protein (eGFP)-tagged L10a ribosomal subunit protein under control of the collagen1α1 promoter. We developed a one-step procedure for isolation of polysomal RNA from collagen1α1-eGFPL10a mice subject to unilateral ureteral obstruction and analyzed and validated the resulting transcriptional profiles. Pathway analysis revealed strong gene signatures for cell proliferation, migration, and shape change. Numerous novel genes and candidate biomarkers were upregulated during fibrosis, specifically in myofibroblasts, and we validated these results by quantitative PCR, in situ, and Western blot analysis. This study provides a comprehensive analysis of early myofibroblast gene expression during kidney fibrosis and introduces a new technique for cell-specific polysomal mRNA isolation in kidney injury models that is suited for RNA-sequencing technologies.
Copyright © 2014 by the American Society of Nephrology.
Figures
References
-
- Schieren G, Rumberger B, Klein M, Kreutz C, Wilpert J, Geyer M, Faller D, Timmer J, Quack I, Rump LC, Walz G, Donauer J: Gene profiling of polycystic kidneys. Nephrol Dial Transplant 21: 1816–1824, 2006 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
