

Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease
- PMID: 24655294
- PMCID: PMC4075043
- DOI: 10.1146/annurev-immunol-032713-120220
Malaria immunity in man and mosquito: insights into unsolved mysteries of a deadly infectious disease
Abstract
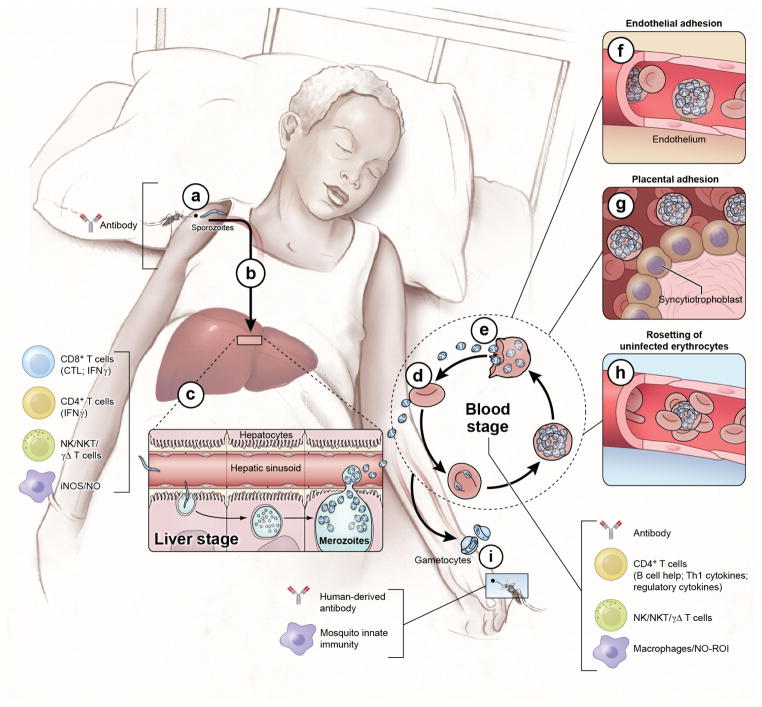
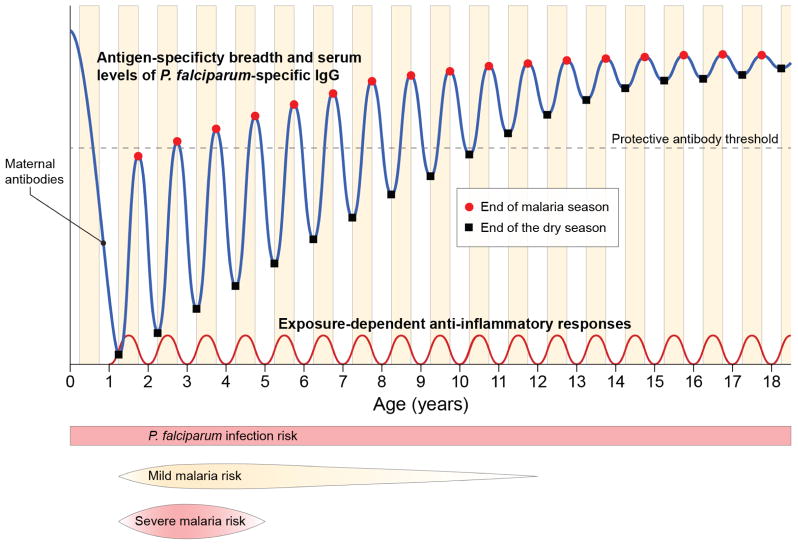
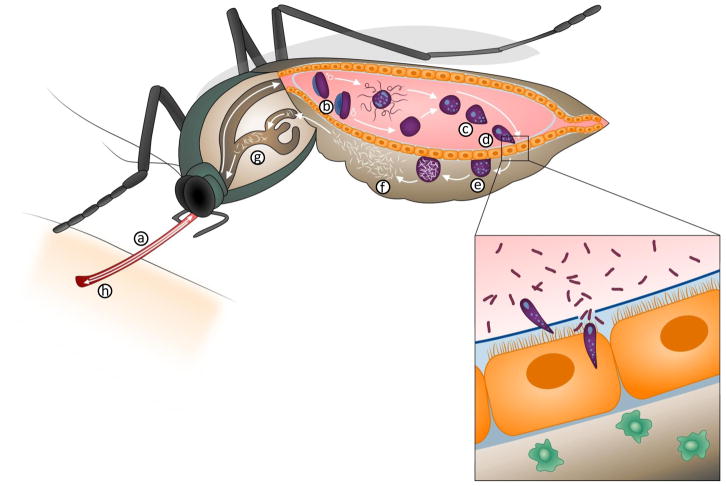
Malaria is a mosquito-borne disease caused by parasites of the obligate intracellular Apicomplexa phylum the most deadly of which, Plasmodium falciparum, prevails in Africa. Malaria imposes a huge health burden on the world's most vulnerable populations, claiming the lives of nearly one million children and pregnant women each year. Although there is keen interest in eradicating malaria, we do not yet have the necessary tools to meet this challenge, including an effective malaria vaccine and adequate vector control strategies. Here we review what is known about the mechanisms at play in immune resistance to malaria in both the human and mosquito hosts at each step in the parasite's complex life cycle with a view toward developing the tools that will contribute to the prevention of disease and death and, ultimately, to the goal of malaria eradication. In so doing, we hope to inspire immunologists to participate in defeating this devastating disease.
Figures
References
-
- Murray CJ, Rosenfeld LC, Lim SS, Andrews KG, Foreman KJ, Haring D, Fullman N, Naghavi M, Lozano R, Lopez AD. Global malaria mortality between 1980 and 2010: a systematic analysis. Lancet. 2012;379:413–31. - PubMed
-
- Langhorne J, Ndungu FM, Sponaas AM, Marsh K. Immunity to malaria: more questions than answers. Nat Immunol. 2008;9:725–32. - PubMed
-
- Marsh K, Kinyanjui S. Immune effector mechanisms in malaria. Parasite Immunol. 2006;28:51–60. - PubMed
-
- Mueller I, Galinski MR, Tsuboi T, Arevalo-Herrera M, Collins WE, King CL. Natural acquisition of immunity to Plasmodium vivax: epidemiological observations and potential targets. Adv Parasitol. 2013;81:77–131. - PubMed
-
- Riley EM, Stewart VA. Immune mechanisms in malaria: new insights in vaccine development. Nat Med. 2013;19:168–78. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
