

Comparative metabolism of cellulose, sophorose and glucose in Trichoderma reesei using high-throughput genomic and proteomic analyses
- PMID: 24655731
- PMCID: PMC3998047
- DOI: 10.1186/1754-6834-7-41
Comparative metabolism of cellulose, sophorose and glucose in Trichoderma reesei using high-throughput genomic and proteomic analyses
Abstract
Background: The filamentous fungus Trichoderma reesei is a major producer of lignocellulolytic enzymes utilized by bioethanol industries. However, to achieve low cost second generation bioethanol production on an industrial scale an efficient mix of hydrolytic enzymes is required for the deconstruction of plant biomass. In this study, we investigated the molecular basis for lignocellulose-degrading enzyme production T. reesei during growth in cellulose, sophorose, and glucose.
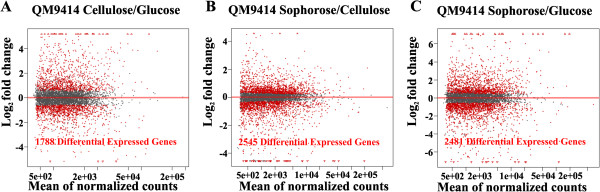
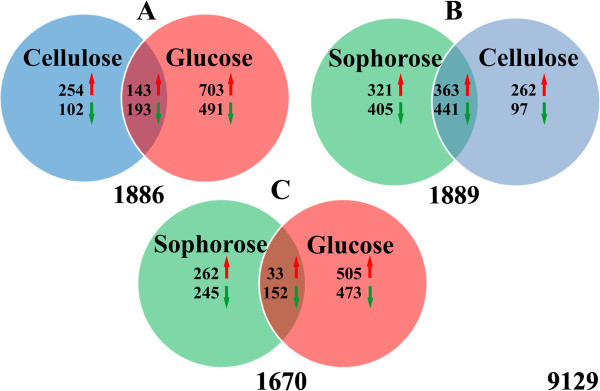
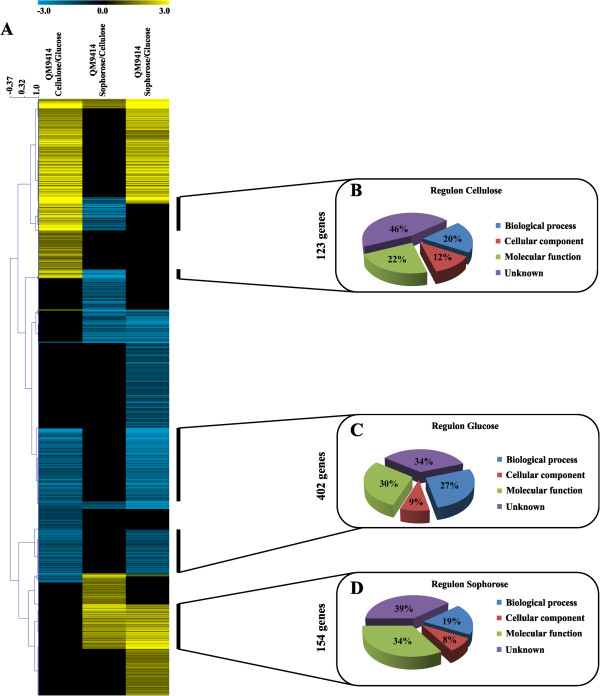
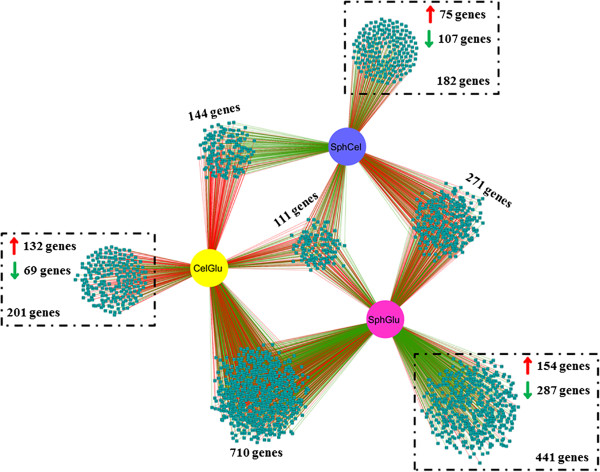
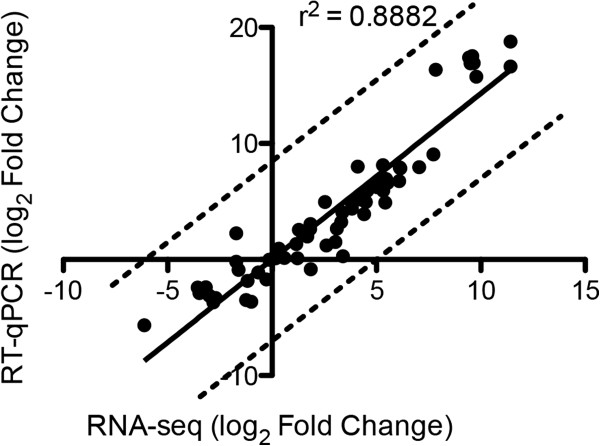
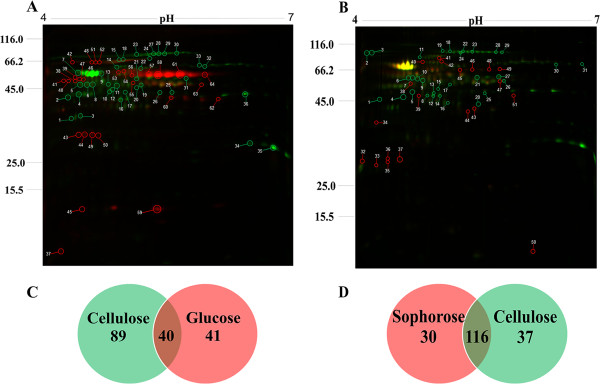
Results: We examined and compared the transcriptome and differential secretome (2D-DIGE) of T. reesei grown in cellulose, sophorose, or glucose as the sole carbon sources. By applying a stringent cut-off threshold 2,060 genes were identified as being differentially expressed in at least one of the respective carbon source comparisons. Hierarchical clustering of the differentially expressed genes identified three possible regulons, representing 123 genes controlled by cellulose, 154 genes controlled by sophorose and 402 genes controlled by glucose. Gene regulatory network analyses of the 692 genes differentially expressed between cellulose and sophorose, identified only 75 and 107 genes as being specific to growth in sophorose and cellulose, respectively. 2D-DIGE analyses identified 30 proteins exclusive to sophorose and 37 exclusive to cellulose. A correlation of 70.17% was obtained between transcription and secreted protein profiles.
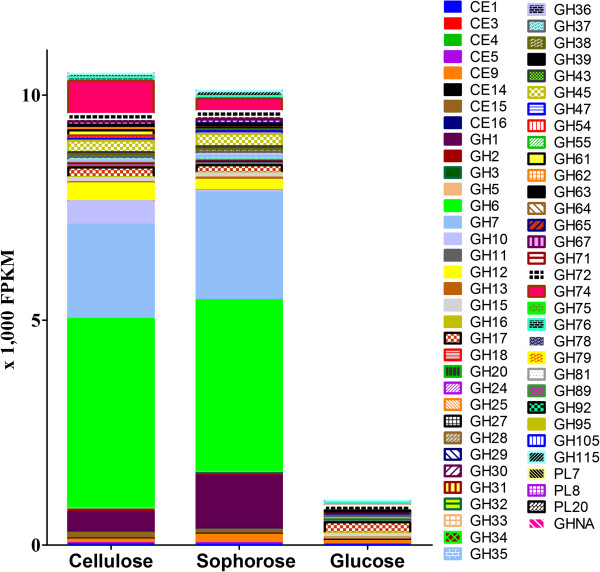
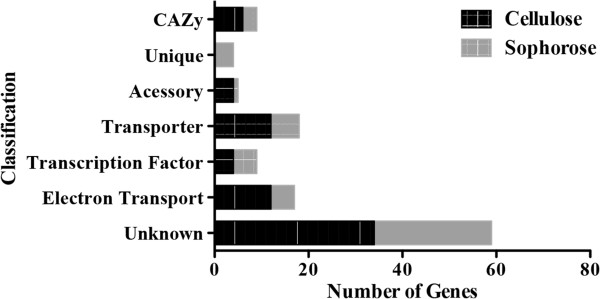
Conclusions: Our data revealed new players in cellulose degradation such as accessory proteins with non-catalytic functions secreted in different carbon sources, transporters, transcription factors, and CAZymes, that specifically respond in response to either cellulose or sophorose.
Figures
References
-
- Pessoa-Jr A, Roberto IC, Menossi M, dos Santos RR, Filho SO, Penna TC. Perspectives on bioenergy and biotechnology in Brazil. Appl Biochem Biotechnol. 2005;121–124:59–70. - PubMed
-
- Ojeda K, Kafarov V. Exergy analysis of enzymatic hydrolysis reactors for transformation of lignocellulosic biomass to bioethanol. Chem Eng J. 2009;154:390–395. doi: 10.1016/j.cej.2009.05.032. - DOI
-
- Soccol CR, Vandenberghe LPD, Medeiros ABP, Karp SG, Buckeridge M, Ramos LP, Pitarelo AP, Ferreira-Leitao V, Gottschalk LMF, Ferrara MA, Bon EPD, de Moraes LMP, Araujo JD, Torres FAG. Bioethanol from lignocelluloses: Status and perspectives in Brazil. Bioresource Technology. 2010;101:4820–4825. doi: 10.1016/j.biortech.2009.11.067. - DOI - PubMed
-
- Kubicek CP, Herrera-Estrella A, Seidl-Seiboth V, Martinez DA, Druzhinina IS, Thon M, Zeilinger S, Casas-Flores S, Horwitz BA, Mukherjee PK, Mukherjee M, Kredics L, Alcaraz LD, Aerts A, Antal Z, Atanasova L, Cervantes-Badillo MG, Challacombe J, Chertkov O, McCluskey K, Coulpier F, Deshpande N, von Dohren H, Ebbole DJ, Esquivel-Naranjo EU, Fekete E, Flipphi M, Glaser F, Gomez-Rodriguez EY, Gruber S. et al.Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome biology. 2011;12:R40. doi: 10.1186/gb-2011-12-4-r40. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
