

Dorsal raphe neurons signal reward through 5-HT and glutamate
- PMID: 24656254
- PMCID: PMC4411946
- DOI: 10.1016/j.neuron.2014.02.010
Dorsal raphe neurons signal reward through 5-HT and glutamate
Abstract
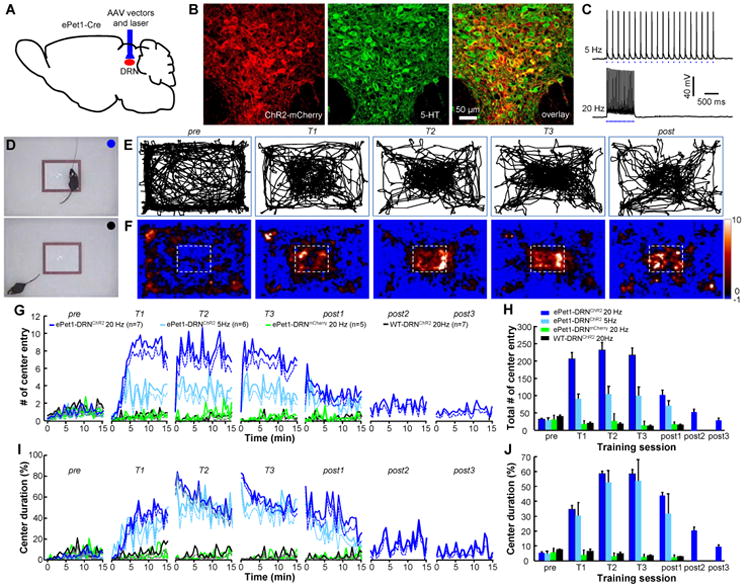
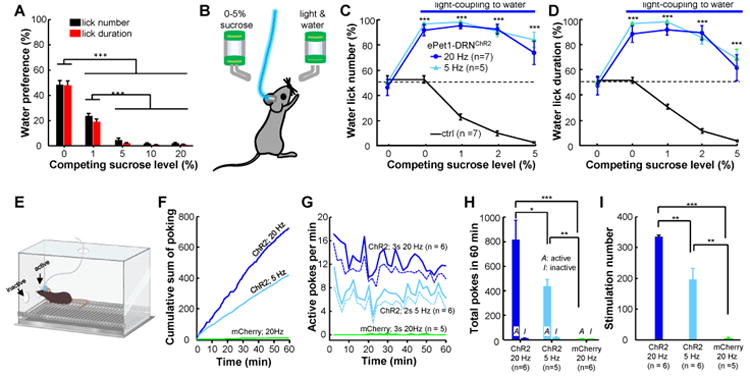
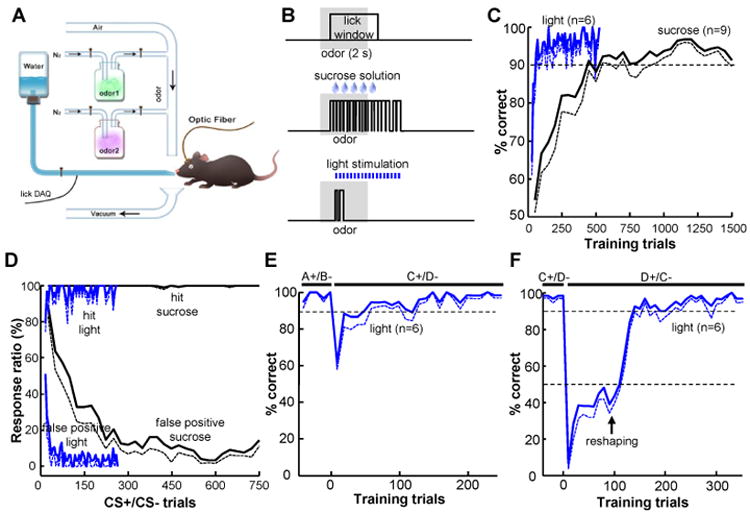
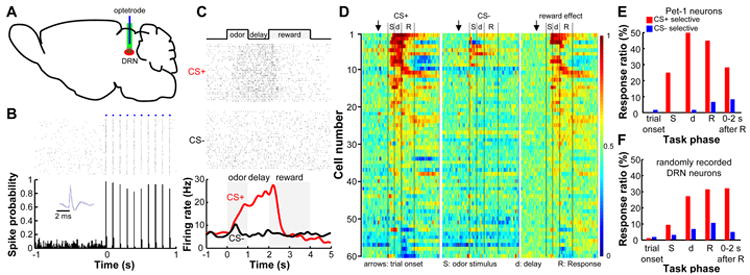
The dorsal raphe nucleus (DRN) in the midbrain is a key center for serotonin (5-hydroxytryptamine; 5-HT)-expressing neurons. Serotonergic neurons in the DRN have been theorized to encode punishment by opposing the reward signaling of dopamine neurons. Here, we show that DRN neurons encode reward, but not punishment, through 5-HT and glutamate. Optogenetic stimulation of DRN Pet-1 neurons reinforces mice to explore the stimulation-coupled spatial region, shifts sucrose preference, drives optical self-stimulation, and directs sensory discrimination learning. DRN Pet-1 neurons increase their firing activity during reward tasks, and this activation can be used to rapidly change neuronal activity patterns in the cortex. Although DRN Pet-1 neurons are often associated with 5-HT, they also release glutamate, and both neurotransmitters contribute to reward signaling. These experiments demonstrate the ability of DRN neurons to organize reward behaviors and might provide insights into the underlying mechanisms of learning facilitation and anhedonia treatment.
Copyright © 2014 Elsevier Inc. All rights reserved.
Figures
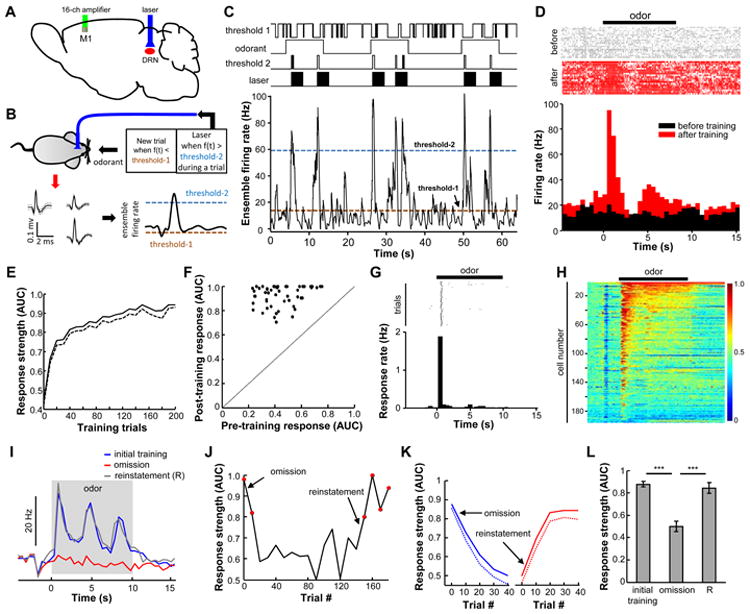
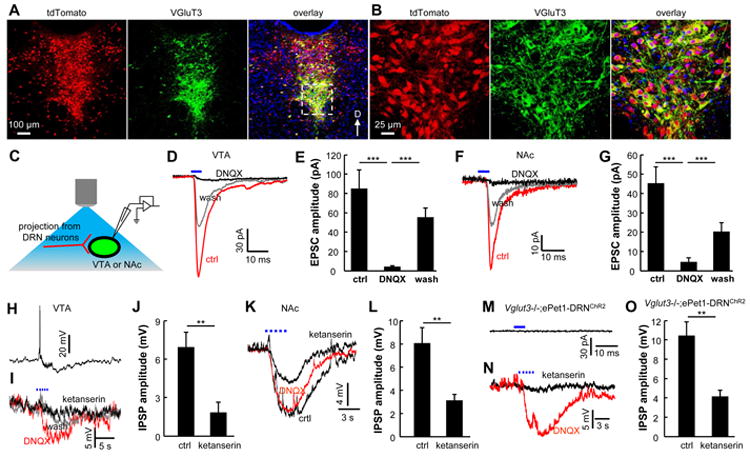
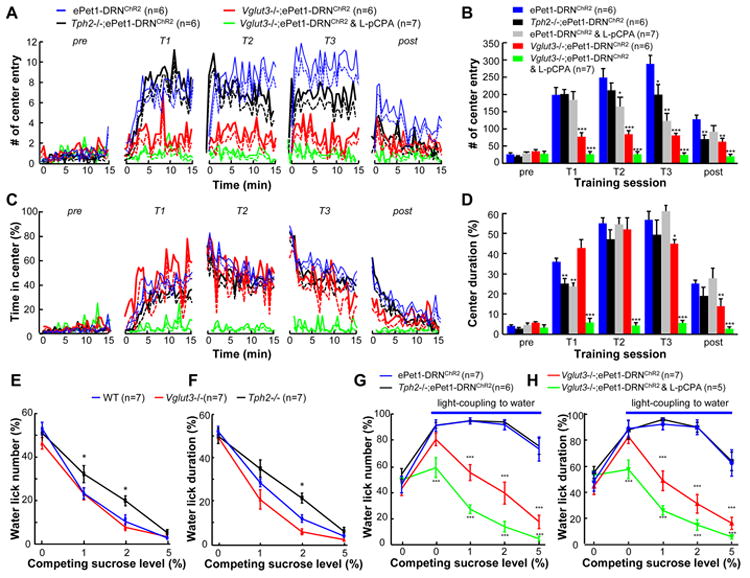
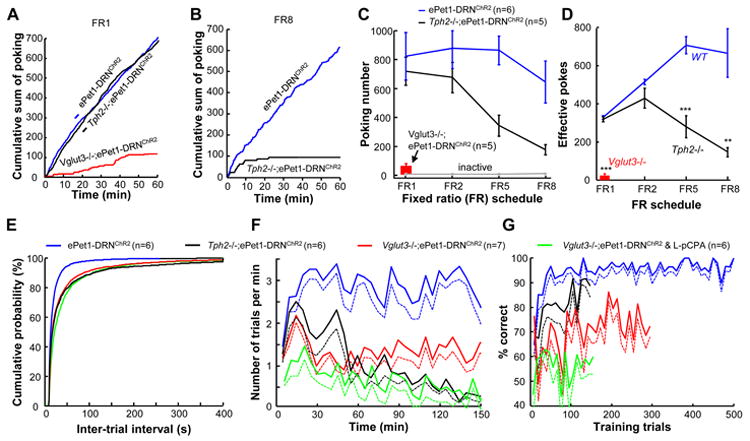
References
-
- Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci. 2005;8:365–371. - PubMed
-
- Amit Z, Smith BR, Gill K. Serotonin uptake inhibitors: effects on motivated consummatory behaviors. J Clin Psychiatry. 1991;52:55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
