

Synthesis and patterning of tunable multiscale materials with engineered cells
- PMID: 24658114
- PMCID: PMC4063449
- DOI: 10.1038/nmat3912
Synthesis and patterning of tunable multiscale materials with engineered cells
Abstract
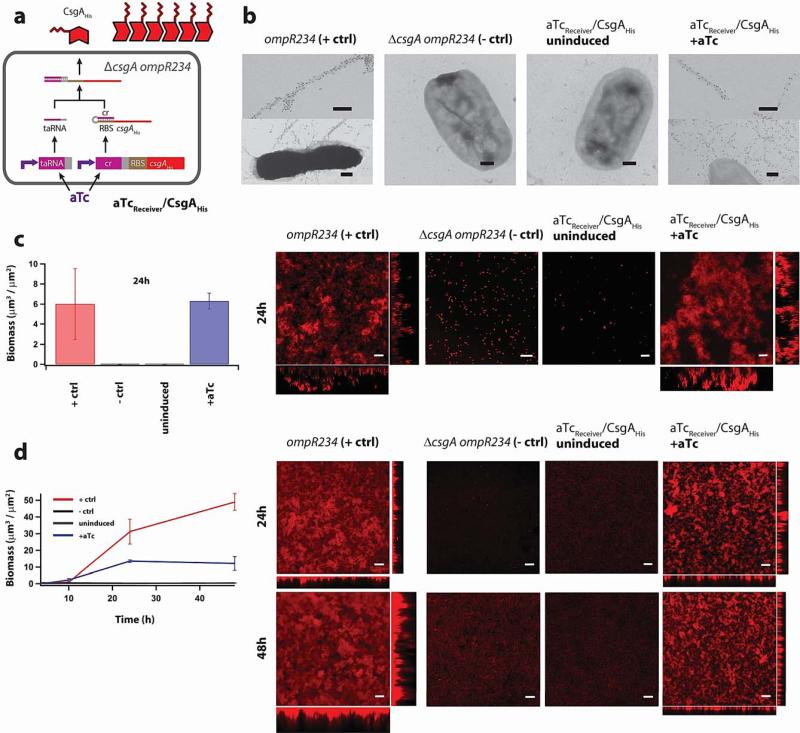
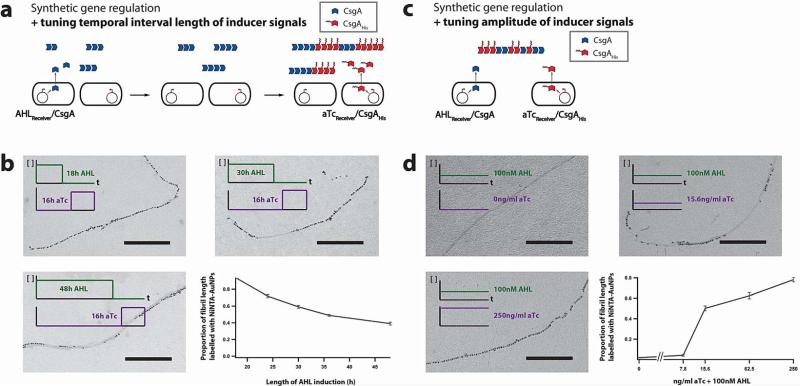
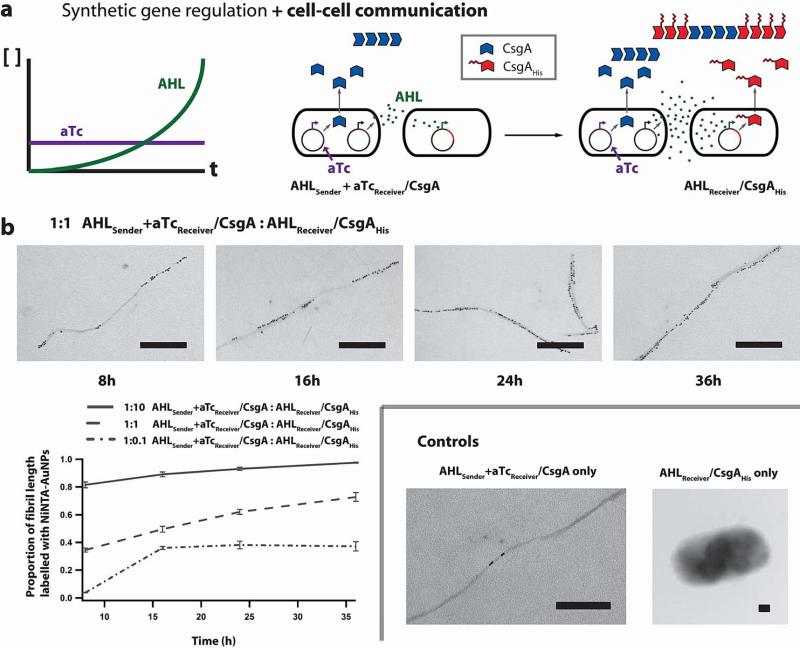
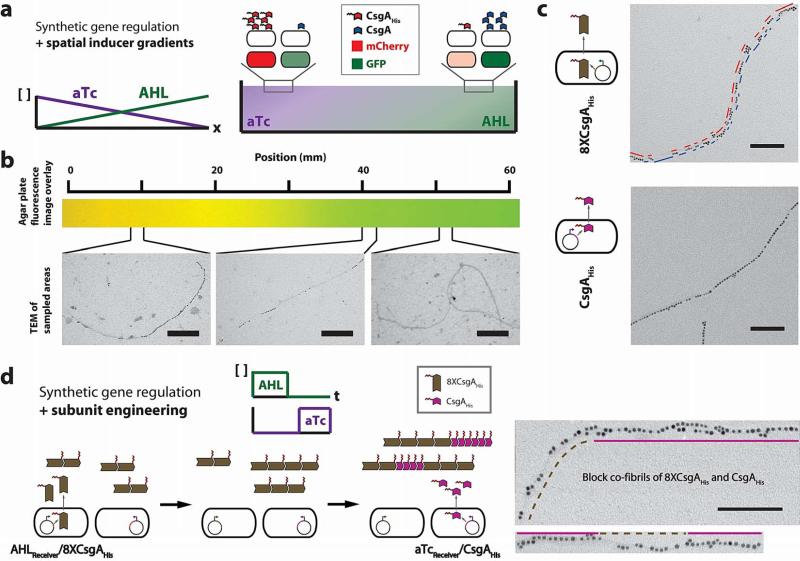
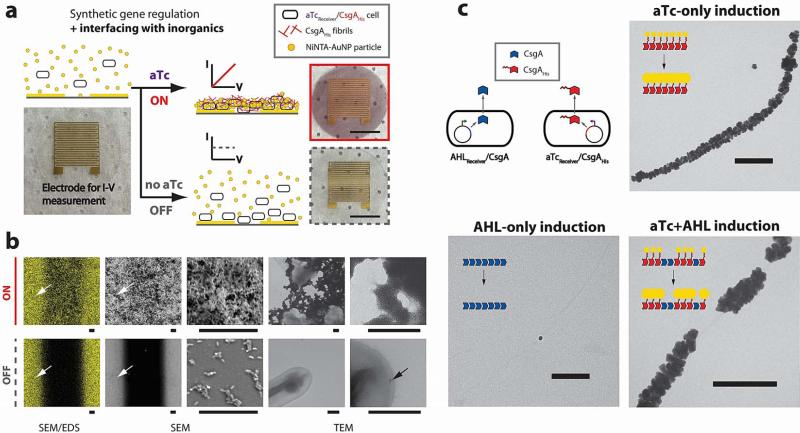
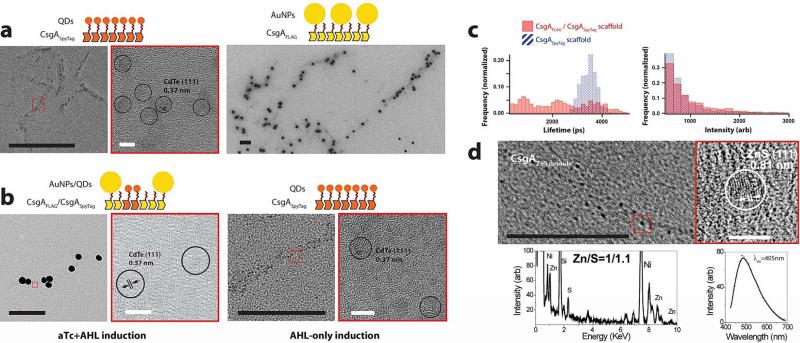
Many natural biological systems--such as biofilms, shells and skeletal tissues--are able to assemble multifunctional and environmentally responsive multiscale assemblies of living and non-living components. Here, by using inducible genetic circuits and cellular communication circuits to regulate Escherichia coli curli amyloid production, we show that E. coli cells can organize self-assembling amyloid fibrils across multiple length scales, producing amyloid-based materials that are either externally controllable or undergo autonomous patterning. We also interfaced curli fibrils with inorganic materials, such as gold nanoparticles (AuNPs) and quantum dots (QDs), and used these capabilities to create an environmentally responsive biofilm-based electrical switch, produce gold nanowires and nanorods, co-localize AuNPs with CdTe/CdS QDs to modulate QD fluorescence lifetimes, and nucleate the formation of fluorescent ZnS QDs. This work lays a foundation for synthesizing, patterning, and controlling functional composite materials with engineered cells.
Figures
References
-
- Fratzl P, Weinkamer R. Nature's hierarchical materials. Progress in Materials Science. 2007;52:1263–1334. doi: http://dx.doi.org/10.1016/j.pmatsci.2007.06.001. - DOI
-
- Kollmannsberger P, Bidan CM, Dunlop JWC, Fratzl P. The physics of tissue patterning and extracellular matrix organisation: how cells join forces. Soft Matter. 2011;7:9549–9560. doi:10.1039/c1sm05588g.
-
- Stevens MM, George JH. Exploring and engineering the cell surface interface. Science. 2005;310:1135–1138. doi:10.1126/science.1106587. - PubMed
-
- O'Toole G, Kaplan HB, Kolter R. Biofilm formation as microbial development. Annu Rev Microbiol. 2000;54:49–79. doi:10.1146/annurev.micro.54.1.4954/1/49 [pii] - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
