

Impaired mitochondrial dynamics and bioenergetics in diabetic skeletal muscle
- PMID: 24658162
- PMCID: PMC3962456
- DOI: 10.1371/journal.pone.0092810
Impaired mitochondrial dynamics and bioenergetics in diabetic skeletal muscle
Erratum in
- PLoS One. 2014;9(6):e101265. LiqunYu [corrected to Yu, Liqun]
Abstract
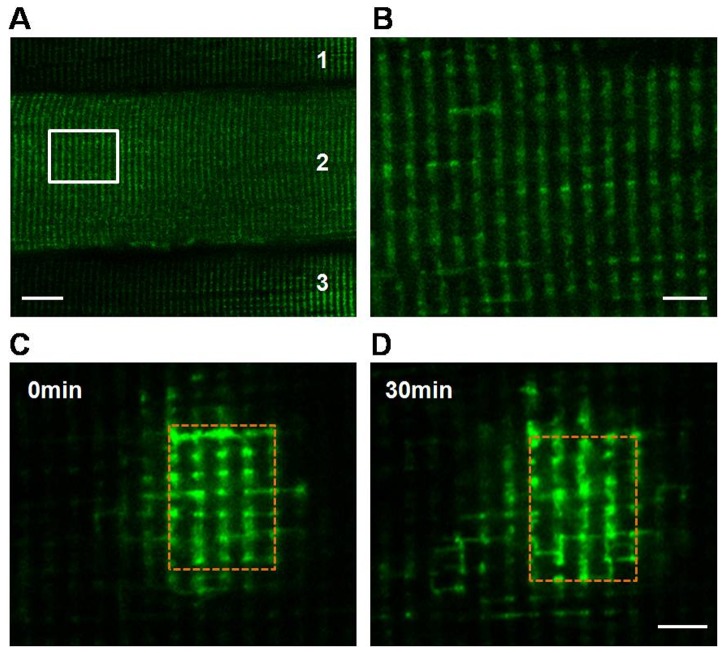
In most cells, mitochondria are highly dynamic organelles that constantly fuse, divide and move. These processes allow mitochondria to redistribute in a cell and exchange contents among the mitochondrial population, and subsequently repair damaged mitochondria. However, most studies on mitochondrial dynamics have been performed on cultured cell lines and neurons, and little is known about whether mitochondria are dynamic organelles in vivo, especially in the highly specialized and differentiated adult skeletal muscle cells. Using mitochondrial matrix-targeted photoactivatable green fluorescent protein (mtPAGFP) and electroporation methods combined with confocal microscopy, we found that mitochondria are dynamic in skeletal muscle in vivo, which enables mitochondria exchange contents within the whole mitochondrial population through nanotunneling-mediated mitochondrial fusion. Mitochondrial network promotes rapid transfer of mtPAGFP within the cell. More importantly, the dynamic behavior was impaired in high-fat diet (HFD)-induced obese mice, accompanying with disturbed mitochondrial respiratory function and decreased ATP content in skeletal muscle. We further found that proteins controlling mitochondrial fusion MFN1 and MFN2 but not Opa1 were decreased and proteins governing mitochondrial fission Fis1 and Drp1 were increased in skeletal muscle of HFD-induced mice when compared to normal diet-fed mice. Altogether, we conclude that mitochondria are dynamic organelles in vivo in skeletal muscle, and it is essential in maintaining mitochondrial respiration and bioenergetics.
Conflict of interest statement
Figures






Similar articles
-
Human skeletal muscle mitochondrial dynamics in relation to oxidative capacity and insulin sensitivity.Diabetologia. 2021 Feb;64(2):424-436. doi: 10.1007/s00125-020-05335-w. Epub 2020 Nov 30. Diabetologia. 2021. PMID: 33258025 Free PMC article.
-
The impact of a short-term high-fat diet on mitochondrial respiration, reactive oxygen species production, and dynamics in oxidative and glycolytic skeletal muscles of young rats.Physiol Rep. 2018 Feb;6(4):e13548. doi: 10.14814/phy2.13548. Physiol Rep. 2018. PMID: 29479852 Free PMC article.
-
Effect of electrical stimulation-induced resistance exercise on mitochondrial fission and fusion proteins in rat skeletal muscle.Appl Physiol Nutr Metab. 2015 Nov;40(11):1137-42. doi: 10.1139/apnm-2015-0184. Epub 2015 Jul 14. Appl Physiol Nutr Metab. 2015. PMID: 26513006
-
Evaluation of in vivo mitochondrial bioenergetics in skeletal muscle using NMR and optical methods.Biochim Biophys Acta. 2016 Apr;1862(4):716-724. doi: 10.1016/j.bbadis.2015.12.019. Epub 2015 Dec 17. Biochim Biophys Acta. 2016. PMID: 26708941 Free PMC article. Review.
-
Implications of mitochondrial fusion and fission in skeletal muscle mass and health.Semin Cell Dev Biol. 2023 Jul 15;143:46-53. doi: 10.1016/j.semcdb.2022.02.011. Epub 2022 Feb 12. Semin Cell Dev Biol. 2023. PMID: 35168898 Review.
Cited by
-
Dysregulated Mitochondrial Dynamics and Metabolism in Obesity, Diabetes, and Cancer.Front Endocrinol (Lausanne). 2019 Sep 3;10:570. doi: 10.3389/fendo.2019.00570. eCollection 2019. Front Endocrinol (Lausanne). 2019. PMID: 31551926 Free PMC article. Review.
-
Mechanisms Orchestrating Mitochondrial Dynamics for Energy Homeostasis.J Mol Biol. 2018 Oct 19;430(21):3922-3941. doi: 10.1016/j.jmb.2018.07.027. Epub 2018 Aug 5. J Mol Biol. 2018. PMID: 30089235 Free PMC article. Review.
-
Influence of Obesity and Type 2 Diabetes on Calcium Handling by Skeletal Muscle: Spotlight on the Sarcoplasmic Reticulum and Mitochondria.Front Physiol. 2021 Nov 2;12:758316. doi: 10.3389/fphys.2021.758316. eCollection 2021. Front Physiol. 2021. PMID: 34795598 Free PMC article. Review.
-
Combining In Vivo 2-Photon Imaging with Photoactivatable Fluorescent Labeling Shows Low Rates of Mitochondrial Dynamics in Skeletal Muscle.Med Sci Sports Exerc. 2025 May 1:10.1249/MSS.0000000000003748. doi: 10.1249/MSS.0000000000003748. Online ahead of print. Med Sci Sports Exerc. 2025. PMID: 40310022 Free PMC article.
-
Autophagy, Innate Immunity and Tissue Repair in Acute Kidney Injury.Int J Mol Sci. 2016 May 3;17(5):662. doi: 10.3390/ijms17050662. Int J Mol Sci. 2016. PMID: 27153058 Free PMC article. Review.
References
-
- Chan DC (2006) Mitochondrial fusion and fission in mammals. Annu Rev Cell Dev Biol 22: 79–99. - PubMed
-
- Westermann B (2010) Mitochondrial fusion and fission in cell life and death. Nat Rev Mol Cell Biol 11: 872–884. - PubMed
-
- Detmer SA, Chan DC (2007) Functions and dysfunctions of mitochondrial dynamics. Nat Rev Mol Cell Biol 8: 870–879. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
