

Genome-wide shRNA screen revealed integrated mitogenic signaling between dopamine receptor D2 (DRD2) and epidermal growth factor receptor (EGFR) in glioblastoma
- PMID: 24658464
- PMCID: PMC4011590
- DOI: 10.18632/oncotarget.1801
Genome-wide shRNA screen revealed integrated mitogenic signaling between dopamine receptor D2 (DRD2) and epidermal growth factor receptor (EGFR) in glioblastoma
Abstract
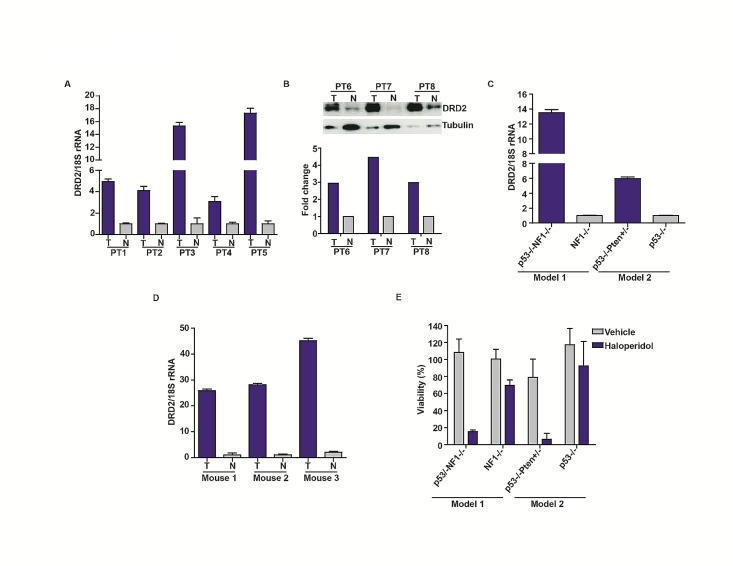
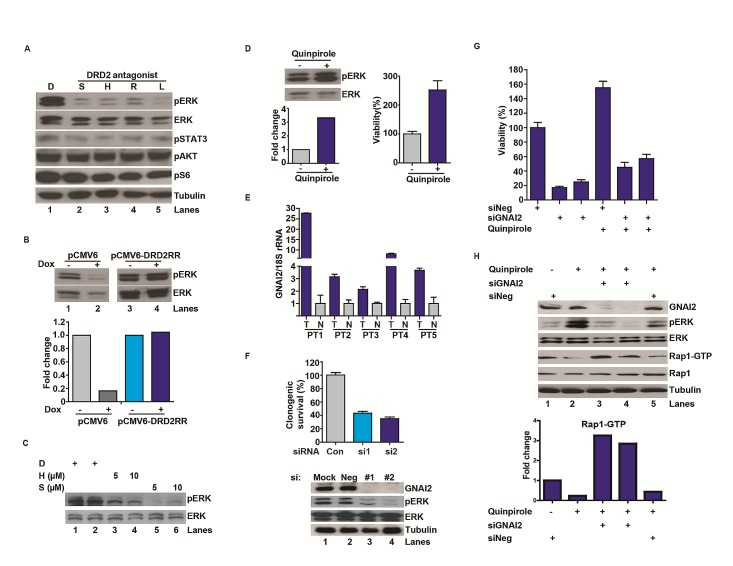
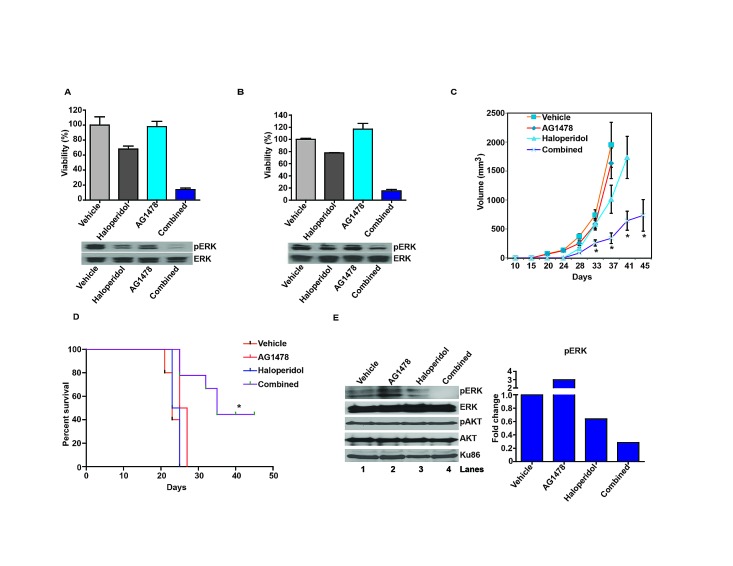
Glioblastoma remains one of the deadliest of human cancers, with most patients succumbing to the disease within two years of diagnosis. The available data suggest that simultaneous inactivation of critical nodes within the glioblastoma molecular circuitry will be required for meaningful clinical efficacy. We conducted parallel genome-wide shRNA screens to identify such nodes and uncovered a number of G-Protein Coupled Receptor (GPCR) neurotransmitter pathways, including the Dopamine Receptor D2 (DRD2) signaling pathway. Supporting the importance of DRD2 in glioblastoma, DRD2 mRNA and protein expression were elevated in clinical glioblastoma specimens relative to matched non-neoplastic cerebrum. Treatment with independent si-/shRNAs against DRD2 or with DRD2 antagonists suppressed the growth of patient-derived glioblastoma lines both in vitro and in vivo. Importantly, glioblastoma lines derived from independent genetically engineered mouse models (GEMMs) were more sensitive to haloperidol, an FDA approved DRD2 antagonist, than the premalignant astrocyte lines by approximately an order of magnitude. The pro-proliferative effect of DRD2 was, in part, mediated through a GNAI2/Rap1/Ras/ERK signaling axis. Combined inhibition of DRD2 and Epidermal Growth Factor Receptor (EGFR) led to synergistic tumoricidal activity as well as ERK suppression in independent in vivo and in vitro glioblastoma models. Our results suggest combined EGFR and DRD2 inhibition as a promising strategy for glioblastoma treatment.
Figures

Comment in
-
Dopamine signaling: target in glioblastoma.Oncotarget. 2014 Mar 15;5(5):1116-7. doi: 10.18632/oncotarget.1835. Oncotarget. 2014. PMID: 24657925 Free PMC article. No abstract available.
References
-
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A, Lacombe D, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352(10):987–996. - PubMed
-
- Wen PY, Kesari S. Malignant gliomas in adults. N Engl J Med. 2008;359(5):492–507. - PubMed
-
- Bartek J, Jr, Ng K, Bartek J, Fischer W, Carter B, Chen CC. Key concepts in glioblastoma therapy. J Neurol Neurosurg Psychiatry. 2012;83(7):753–760. - PubMed
-
- Ng K, Kim R, Kesari S, Carter B, Chen CC. Genomic profiling of glioblastoma: convergence of fundamental biologic tenets and novel insights. J Neurooncol. 2012;107(1):1–12. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
