

Integrin α4β1 is necessary for CD4+ T cell-mediated protection against genital Chlamydia trachomatis infection
- PMID: 24659687
- PMCID: PMC3995848
- DOI: 10.4049/jimmunol.1303238
Integrin α4β1 is necessary for CD4+ T cell-mediated protection against genital Chlamydia trachomatis infection
Abstract
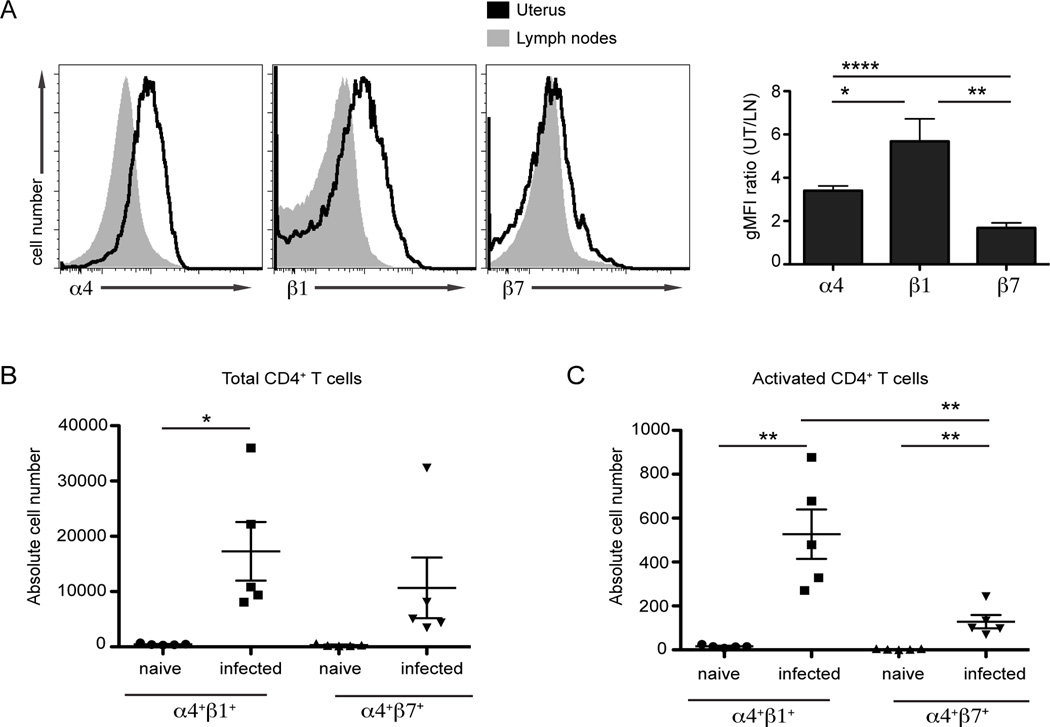
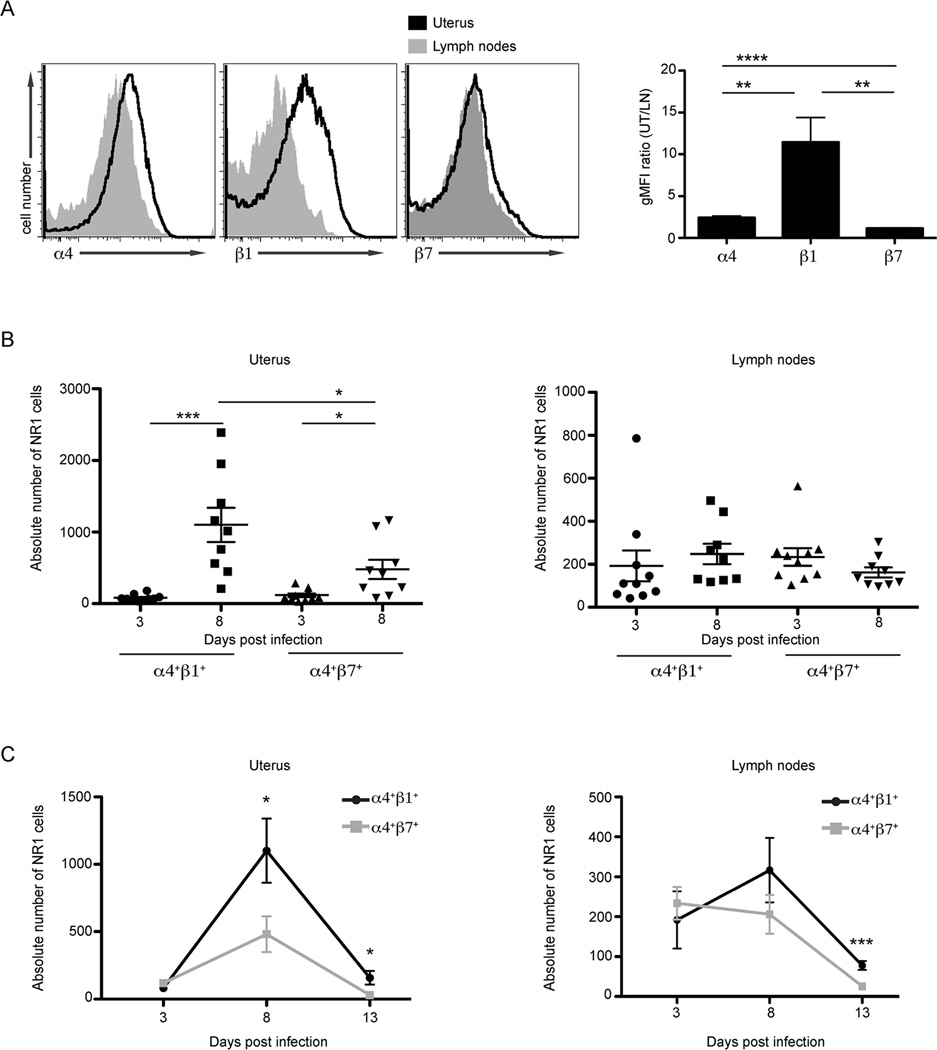
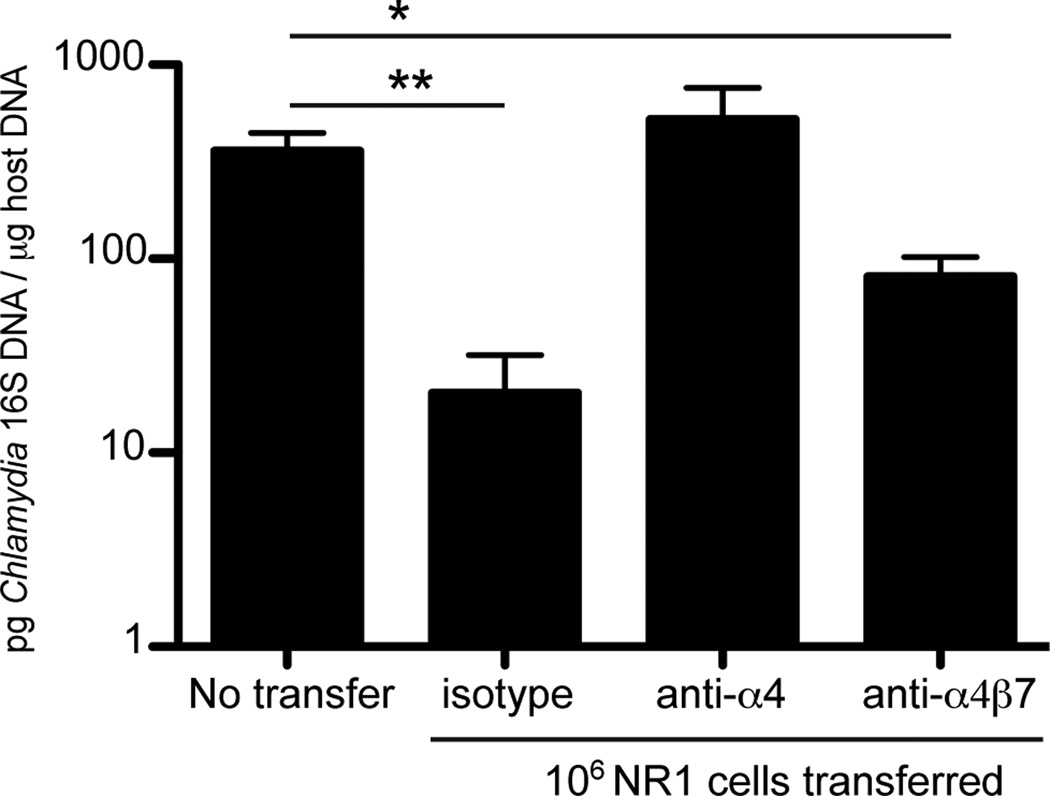
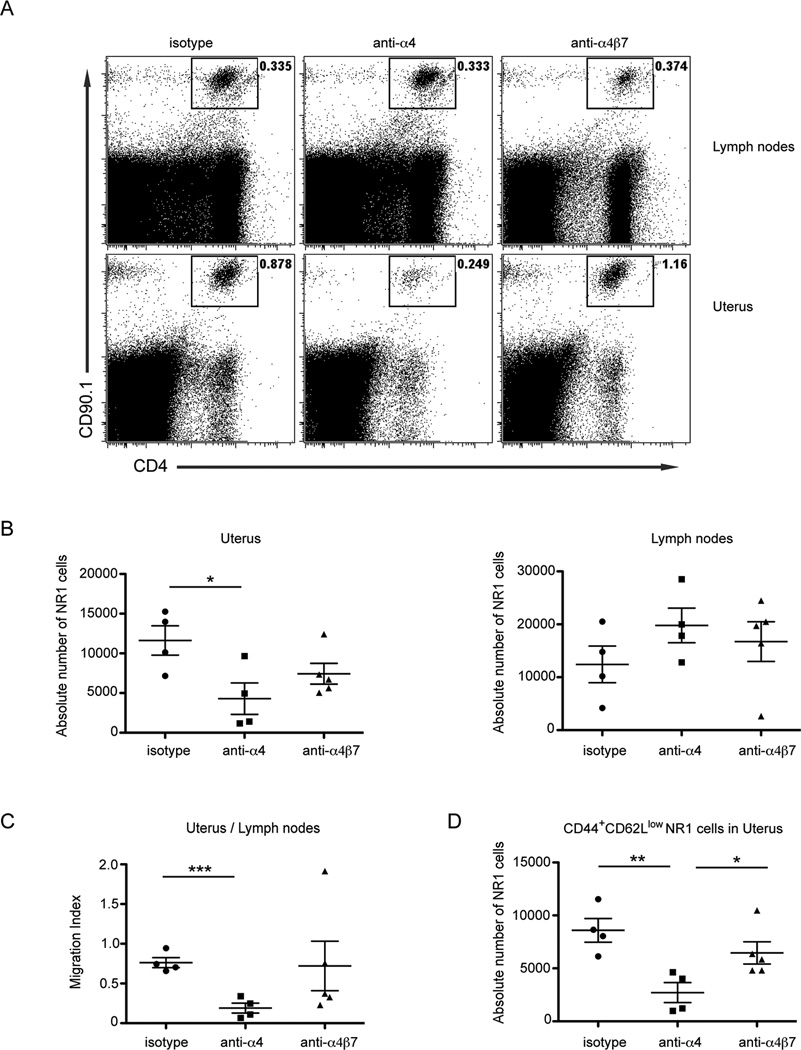
Chlamydia trachomatis infection is the most common sexually transmitted bacterial infection in the United States and a significant health burden worldwide. Protection from Chlamydia infection in the genital mucosa is dependent on IFN-γ derived from CD4(+) Th1 cells. These CD4(+) T cells must home successfully to the genital tract to exert their effector function and decrease C. trachomatis burden. Although adhesion receptors expressed by CD4(+) T cells in the genital tract have been characterized, the integrin receptor required for Chlamydia-specific CD4(+) T cell-mediated protection has not been explored. In this study, we demonstrate that C. trachomatis infection of the upper genital tract results in recruitment of Chlamydia-specific CD4(+) T cells robustly expressing the integrin α4β1. Interfering with α4β1, but not α4β7, function resulted in defective CD4(+) T cell trafficking to the uterus and high bacterial load. We conclude that integrin α4β1 is necessary for CD4(+) T cell-mediated protection against C. trachomatis infection in the genital mucosa. By identifying homing molecules required for successful CD4(+) T cell trafficking to C. trachomatis-infected tissues, we will be better equipped to design vaccines that elicit sterilizing, long-lasting immunity without inducing immune pathologies in the upper genital tract.
Conflict of interest statement
The authors declare no financial conflicts of interest.
Figures


References
-
- Brunham R, Rey-Ladino J. Immunology of Chlamydia infection: implications for a Chlamydia trachomatis vaccine. Nature reviews. Immunology. 2005;5:149–161. - PubMed
-
- Roan N, Starnbach M. Immune-mediated control of Chlamydia infection. Cellular microbiology. 2008;10:9–19. - PubMed
-
- Perry L, Su H, Feilzer K, Messer R, Hughes S, Whitmire W, Caldwell H. Differential sensitivity of distinct Chlamydia trachomatis isolates to IFN-gamma-mediated inhibition. Journal of immunology (Baltimore, Md. : 1950) 1999;162:3541–3548. - PubMed
-
- Thomas S, Garrity L, Brandt C, Schobert C, Feng G, Taylor M, Carlin J, Byrne G. IFN-gamma-mediated antimicrobial response. Indoleamine 2,3-dioxygenase-deficient mutant host cells no longer inhibit intracellular Chlamydia spp. or Toxoplasma growth. Journal of immunology (Baltimore, Md. : 1950) 1993;150:5529–5534. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
