

Intracellular hydrogen peroxide and superoxide poison 3-deoxy-D-arabinoheptulosonate 7-phosphate synthase, the first committed enzyme in the aromatic biosynthetic pathway of Escherichia coli
- PMID: 24659765
- PMCID: PMC4010980
- DOI: 10.1128/JB.01573-14
Intracellular hydrogen peroxide and superoxide poison 3-deoxy-D-arabinoheptulosonate 7-phosphate synthase, the first committed enzyme in the aromatic biosynthetic pathway of Escherichia coli
Abstract
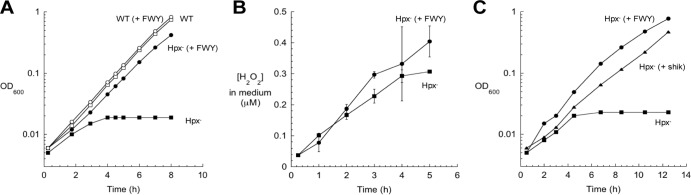
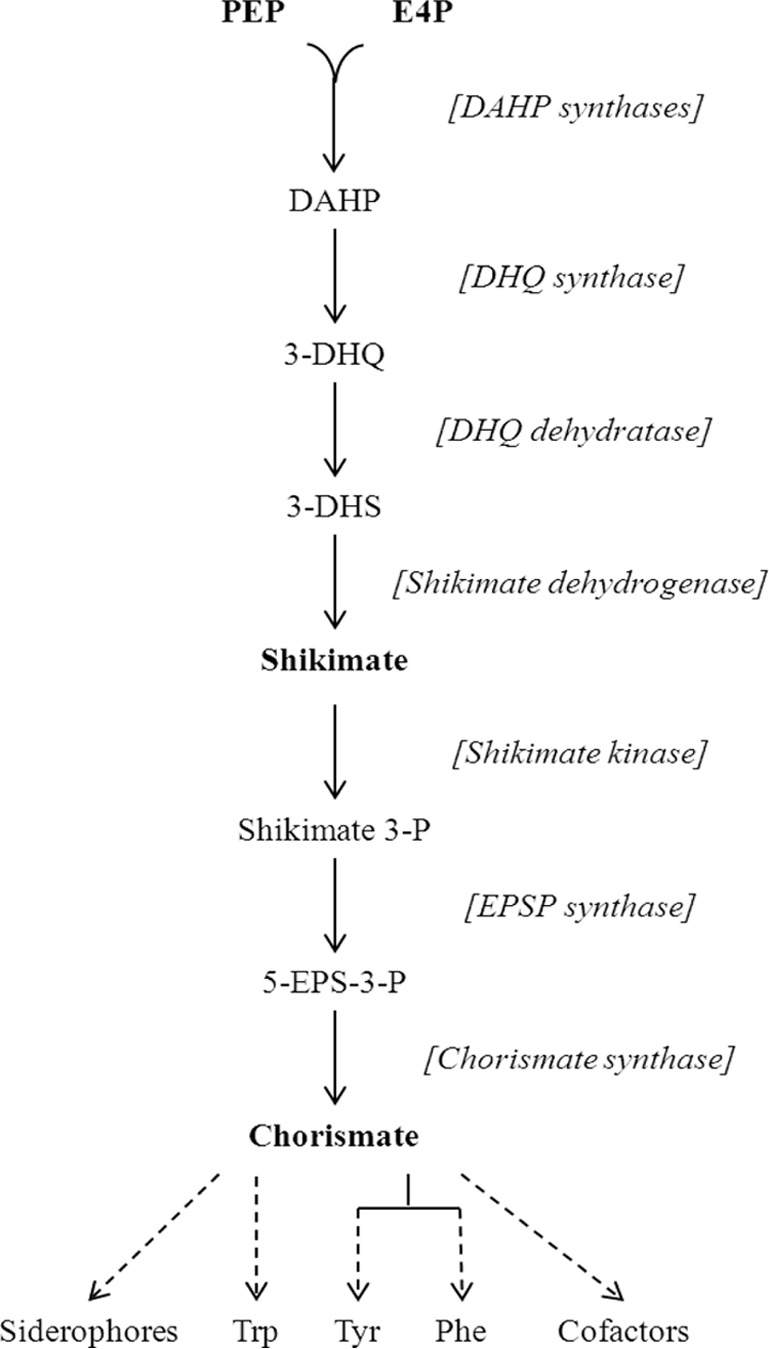
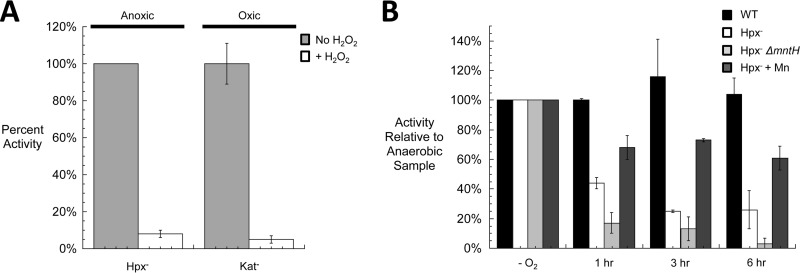
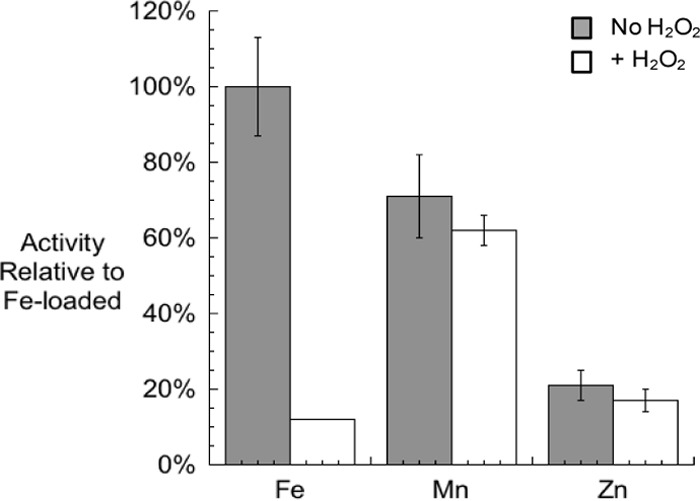
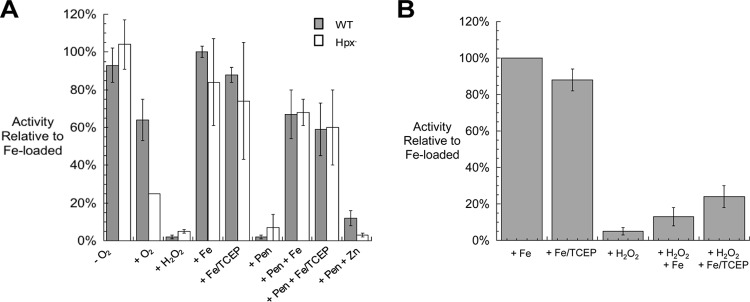
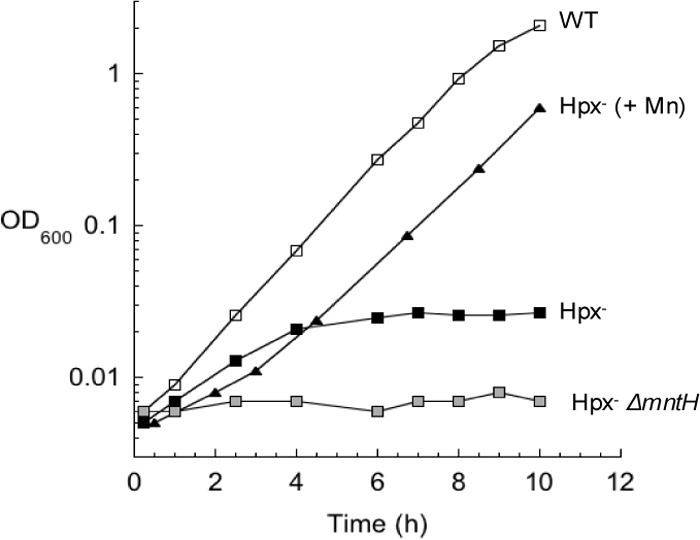
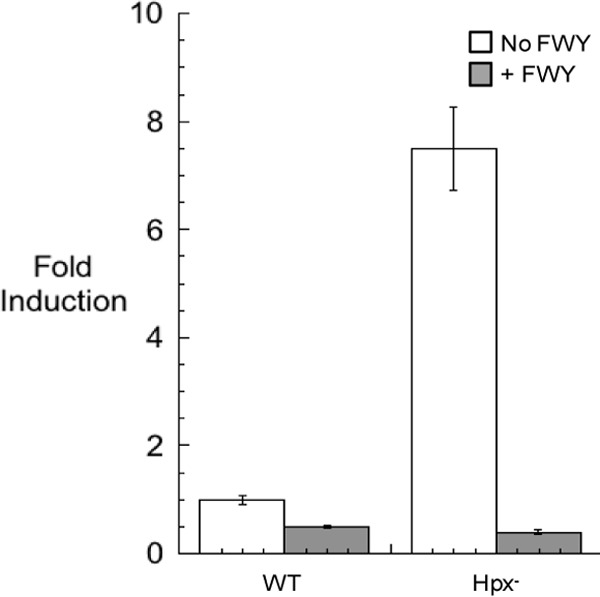
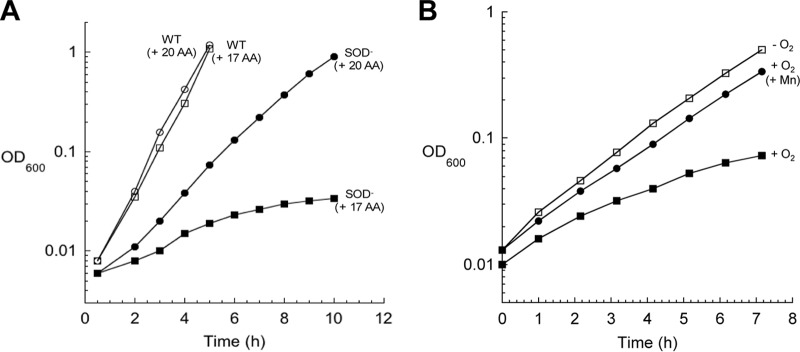
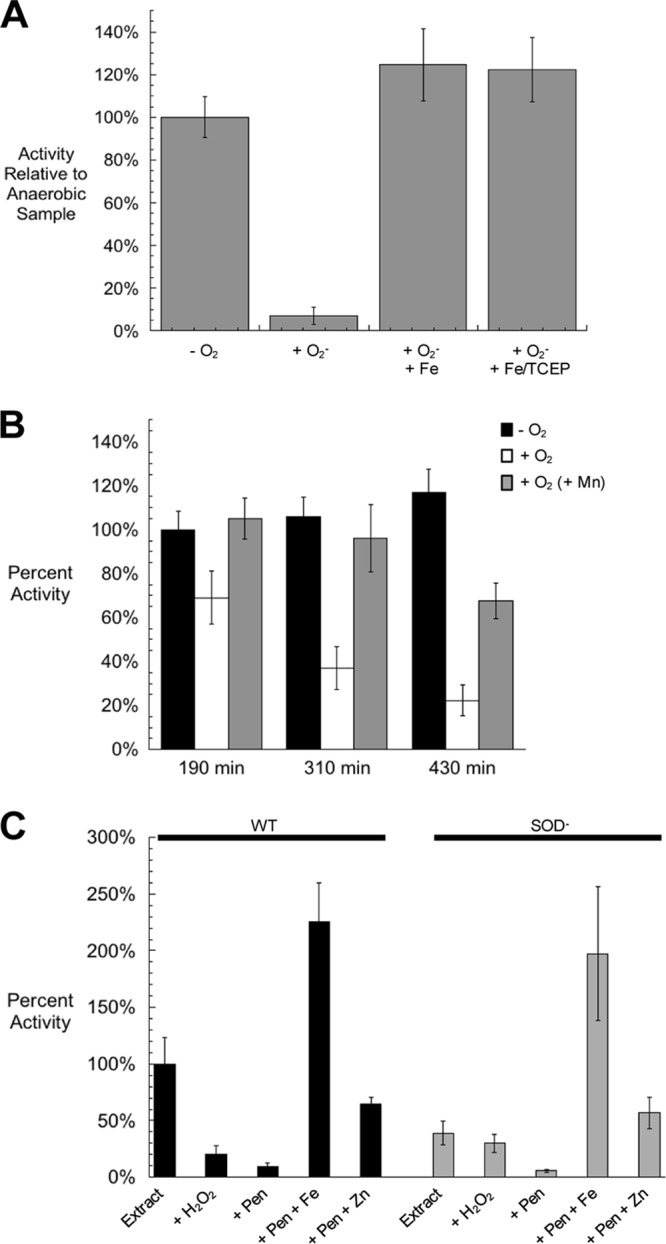
In Escherichia coli, aromatic compound biosynthesis is the process that has shown the greatest sensitivity to hydrogen peroxide stress. This pathway has long been recognized to be sensitive to superoxide as well, but the molecular target was unknown. Feeding experiments indicated that the bottleneck lies early in the pathway, and the suppressive effects of fur mutations and manganese supplementation suggested the involvement of a metalloprotein. The 3-deoxy-D-arabinoheptulosonate 7-phosphate synthase (DAHP synthase) activity catalyzes the first step in the pathway, and it is provided by three isozymes known to rely upon a divalent metal. This activity progressively declined when cells were stressed with either oxidant. The purified enzyme was activated more strongly by ferrous iron than by other metals, and only this metalloform could be inactivated by hydrogen peroxide or superoxide. We infer that iron is the prosthetic metal in vivo. Both oxidants displace the iron atom from the enzyme. In peroxide-stressed cells, the enzyme accumulated as an apoprotein, potentially with an oxidized cysteine residue. In superoxide-stressed cells, the enzyme acquired a nonactivating zinc ion in its active site, an apparent consequence of the repeated ejection of iron. Manganese supplementation protected the activity in both cases, which matches the ability of manganese to metallate the enzyme and to provide substantial oxidant-resistant activity. DAHP synthase thus belongs to a family of mononuclear iron-containing enzymes that are disabled by oxidative stress. To date, all the intracellular injuries caused by physiological doses of these reactive oxygen species have arisen from the oxidation of reduced iron centers.
Figures
References
-
- McCord JM, Fridovich I. 1969. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 244:6049–6055 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
