

Establishment of chronic hepatitis C virus infection: translational evasion of oxidative defence
- PMID: 24659872
- PMCID: PMC3961964
- DOI: 10.3748/wjg.v20.i11.2785
Establishment of chronic hepatitis C virus infection: translational evasion of oxidative defence
Abstract
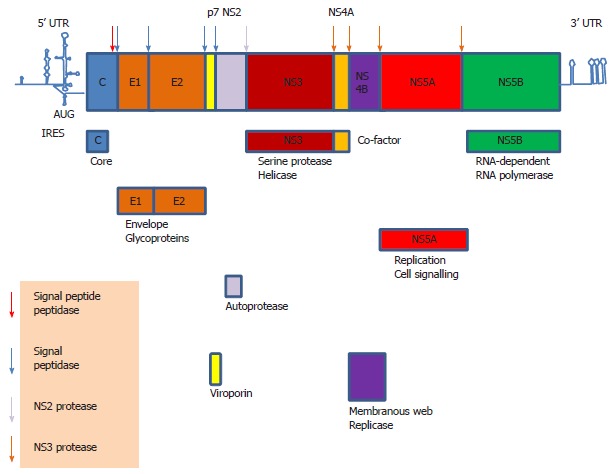
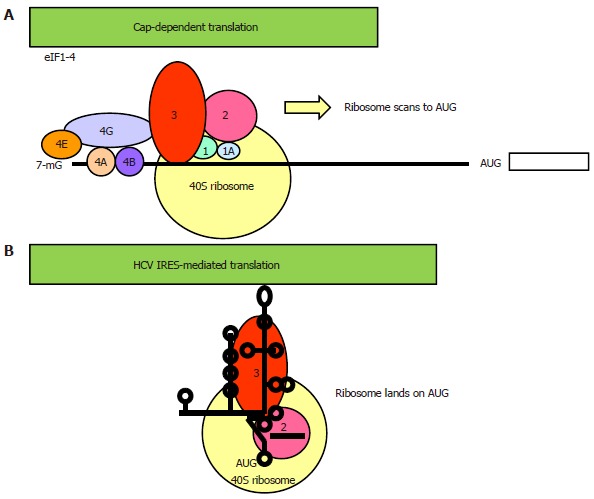
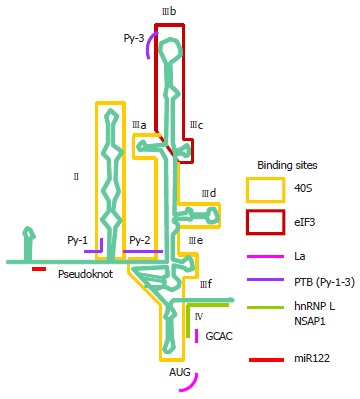
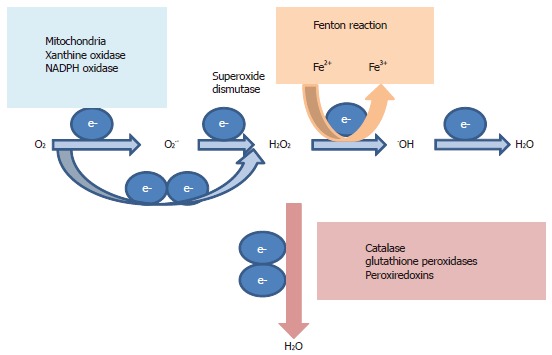
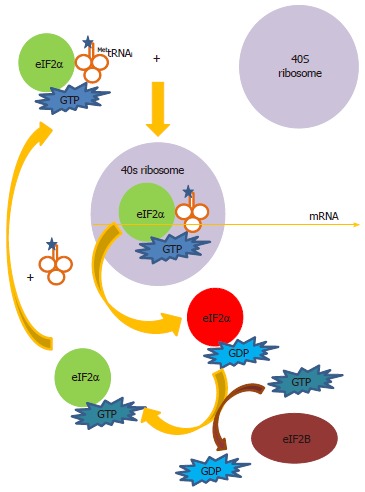
Hepatitis C virus (HCV) causes a clinically important disease affecting 3% of the world population. HCV is a single-stranded, positive-sense RNA virus belonging to the genus Hepacivirus within the Flaviviridae family. The virus establishes a chronic infection in the face of an active host oxidative defence, thus adaptation to oxidative stress is key to virus survival. Being a small RNA virus with a limited genomic capacity, we speculate that HCV deploys a different strategy to evade host oxidative defence. Instead of counteracting oxidative stress, it utilizes oxidative stress to facilitate its own survival. Translation is the first step in the replication of a plus strand RNA virus so it would make sense if the virus can exploit the host oxidative defence in facilitating this very first step. This is particularly true when HCV utilizes an internal ribosome entry site element in translation, which is distinctive from that of cap-dependent translation of the vast majority of cellular genes, thus allowing selective translation of genes under conditions when global protein synthesis is compromised. Indeed, we were the first to show that HCV translation was stimulated by an important pro-oxidant-hydrogen peroxide in hepatocytes, suggesting that HCV is able to adapt to and utilize the host anti-viral response to facilitate its own translation thus allowing the virus to thrive under oxidative stress condition to establish chronicity. Understanding how HCV translation is regulated under oxidative stress condition will advance our knowledge on how HCV establishes chronicity. As chronicity is the initiator step in disease progression this will eventually lead to a better understanding of pathogenicity, which is particularly relevant to the development of anti-virals and improved treatments of HCV patients using anti-oxidants.
Keywords: Chronicity; Hepatitis C virus; Hydrogen peroxide; Internal ribosome entry site; Oxidative stress; Persistence; Translation.
Figures
References
-
- Hajarizadeh B, Grebely J, Dore GJ. Epidemiology and natural history of HCV infection. Nat Rev Gastroenterol Hepatol. 2013;10:553–562. - PubMed
-
- Bowen DG, Walker CM. Adaptive immune responses in acute and chronic hepatitis C virus infection. Nature. 2005;436:946–952. - PubMed
-
- Gale M, Foy EM. Evasion of intracellular host defence by hepatitis C virus. Nature. 2005;436:939–945. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
