

Interleukin-6 modulation of intestinal epithelial tight junction permeability is mediated by JNK pathway activation of claudin-2 gene
- PMID: 24662742
- PMCID: PMC3963839
- DOI: 10.1371/journal.pone.0085345
Interleukin-6 modulation of intestinal epithelial tight junction permeability is mediated by JNK pathway activation of claudin-2 gene
Abstract
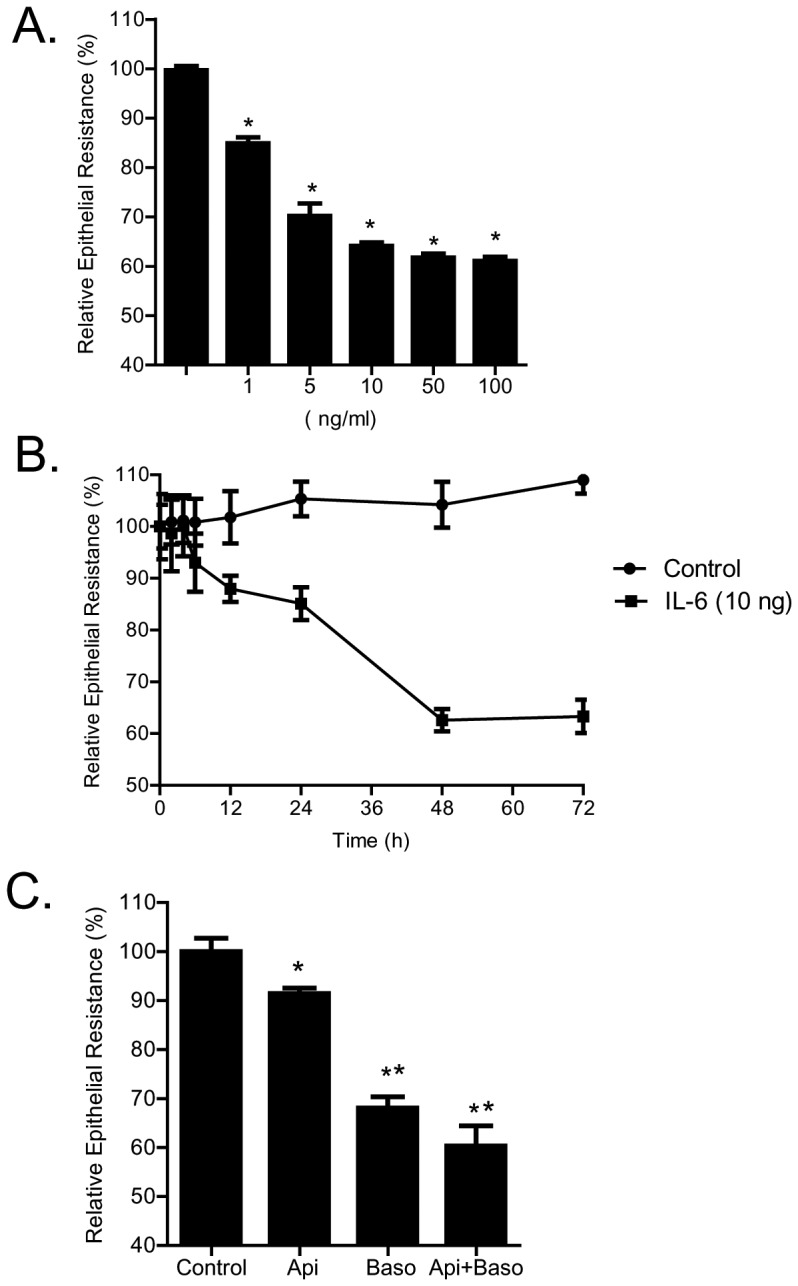
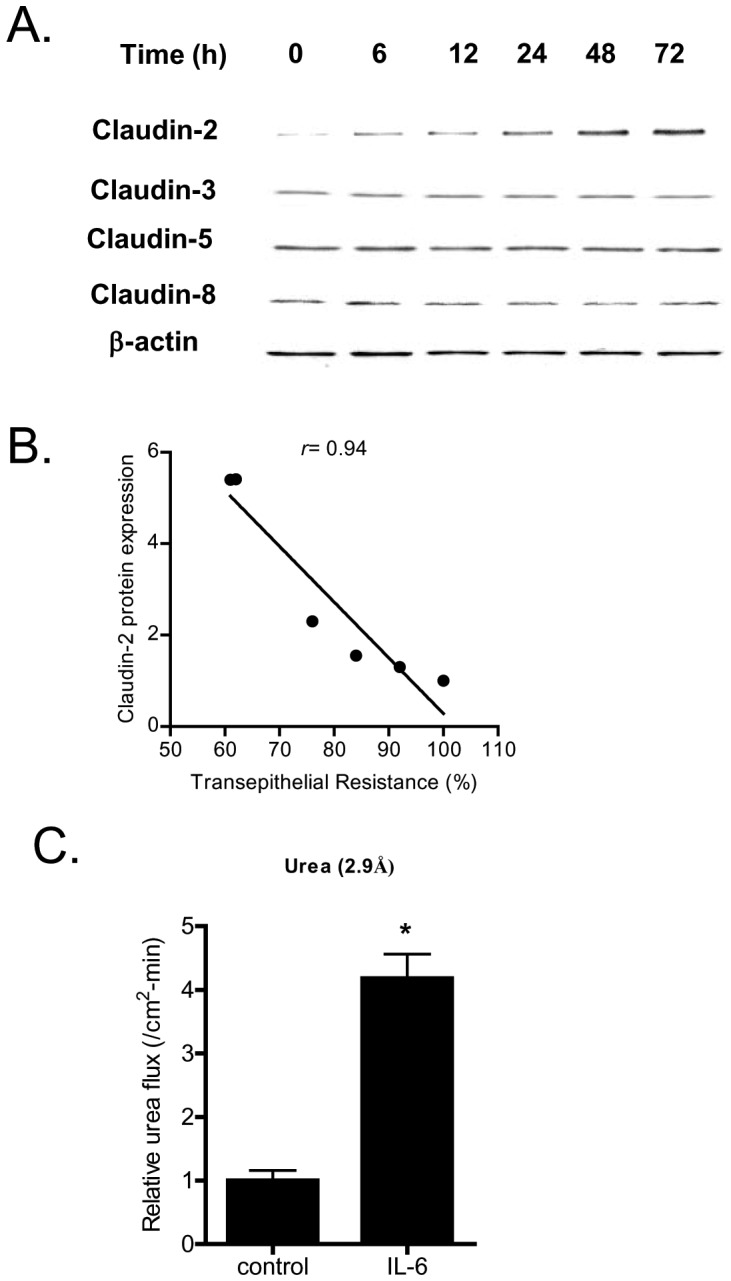
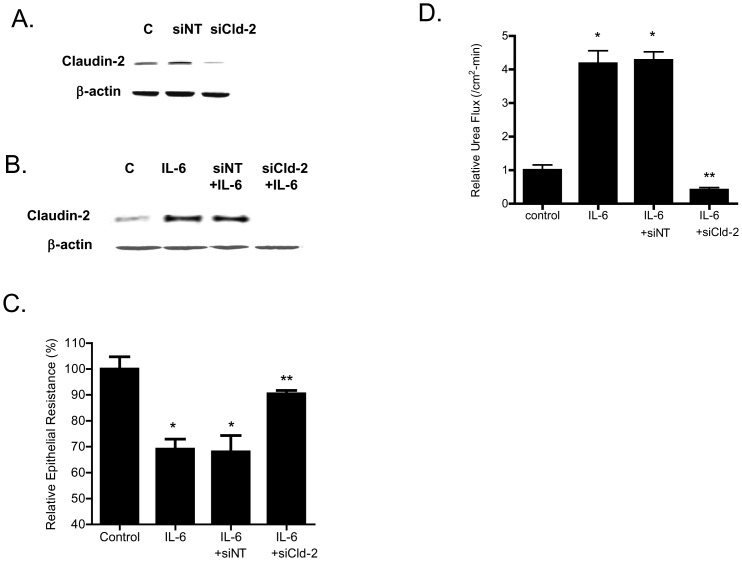
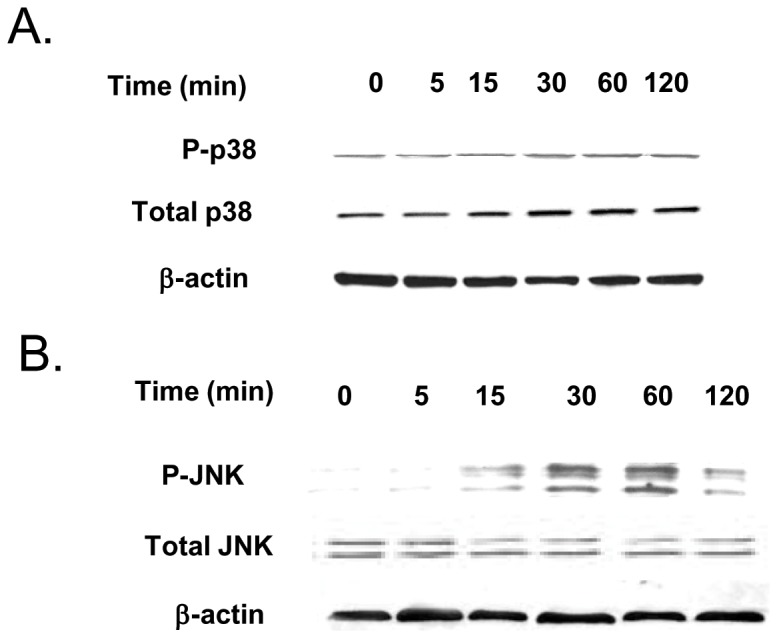
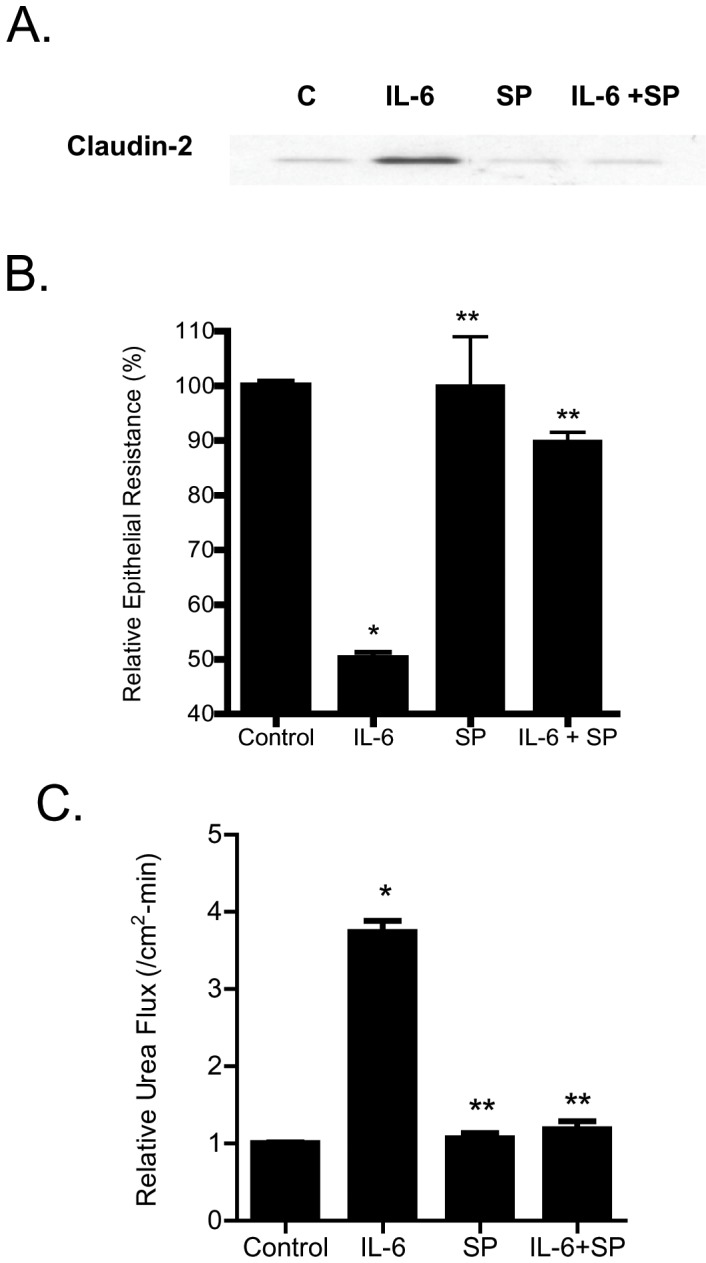
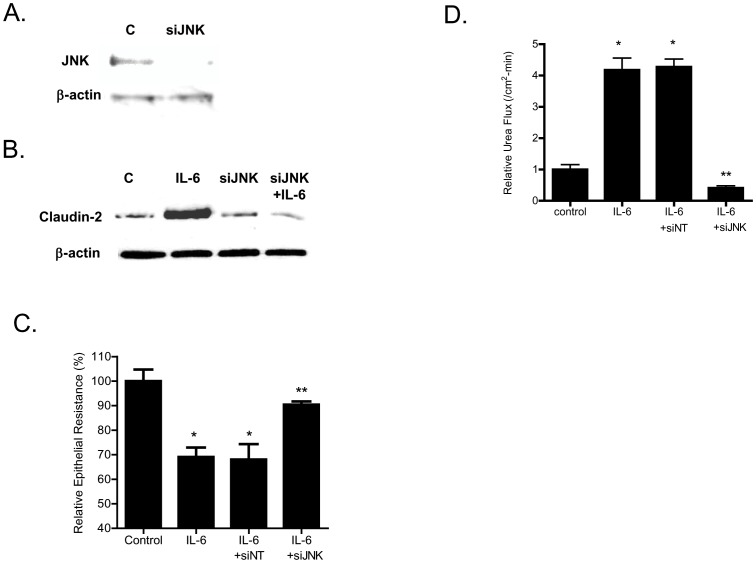
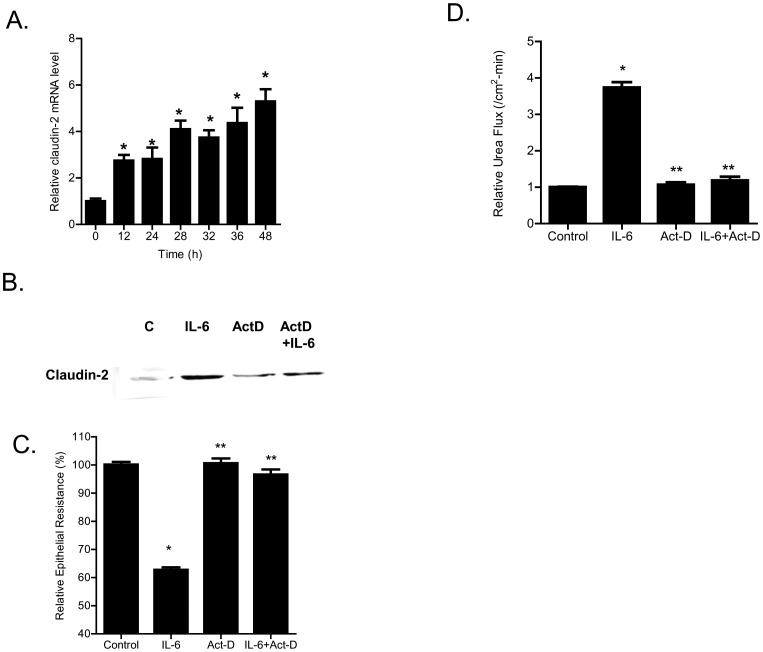
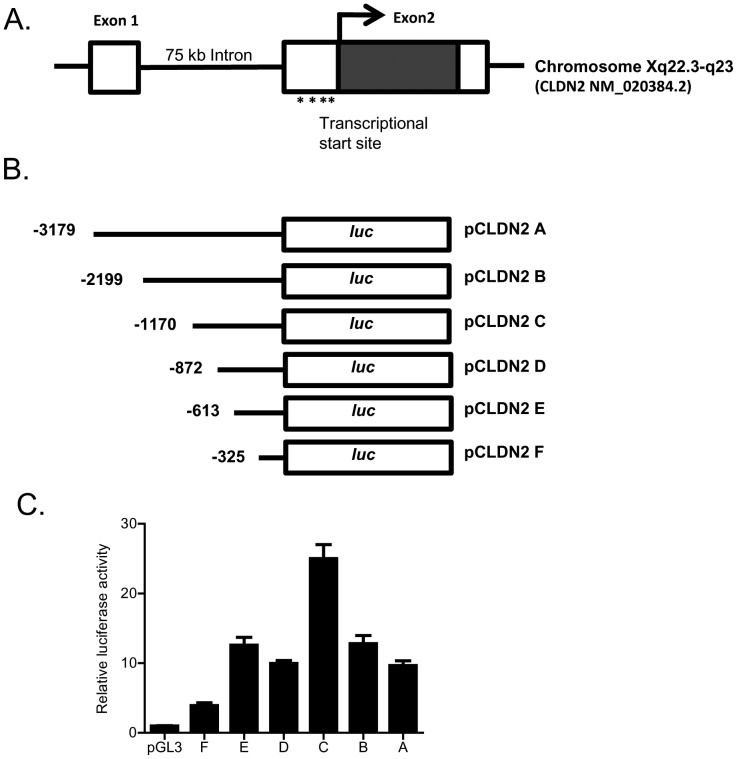
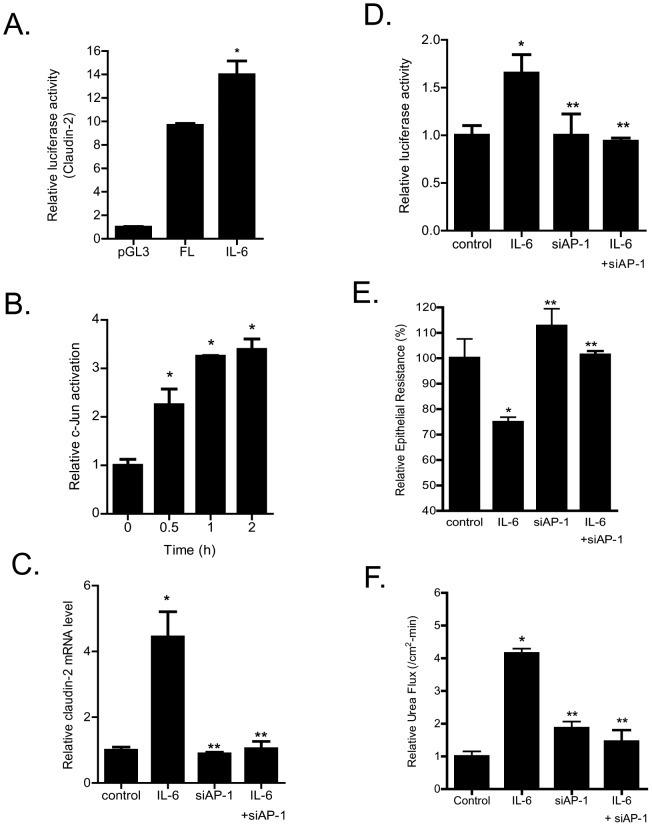
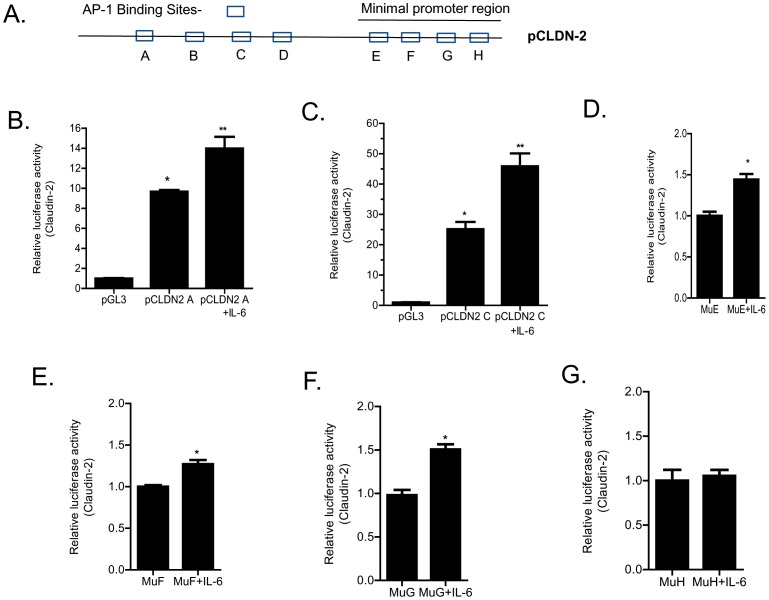
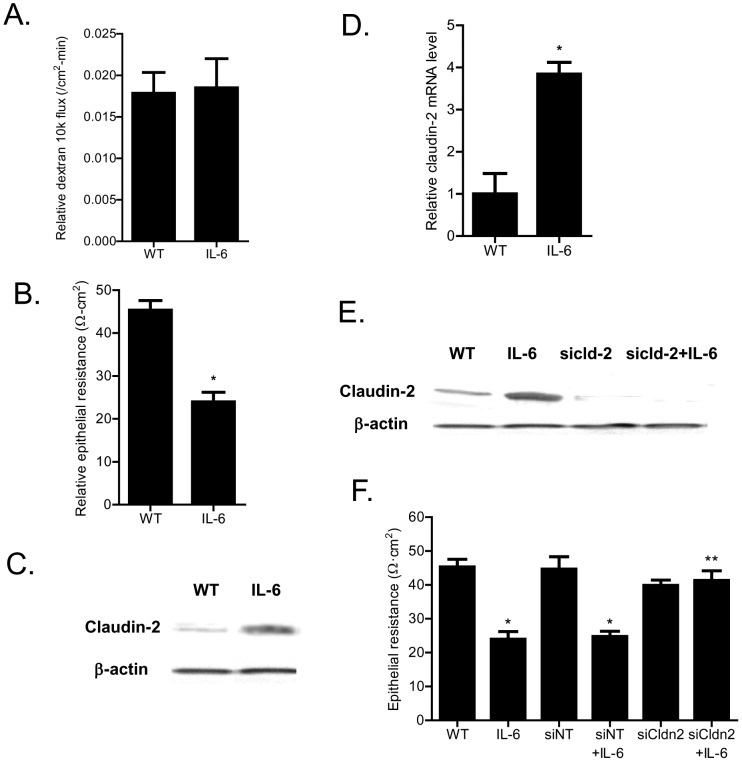
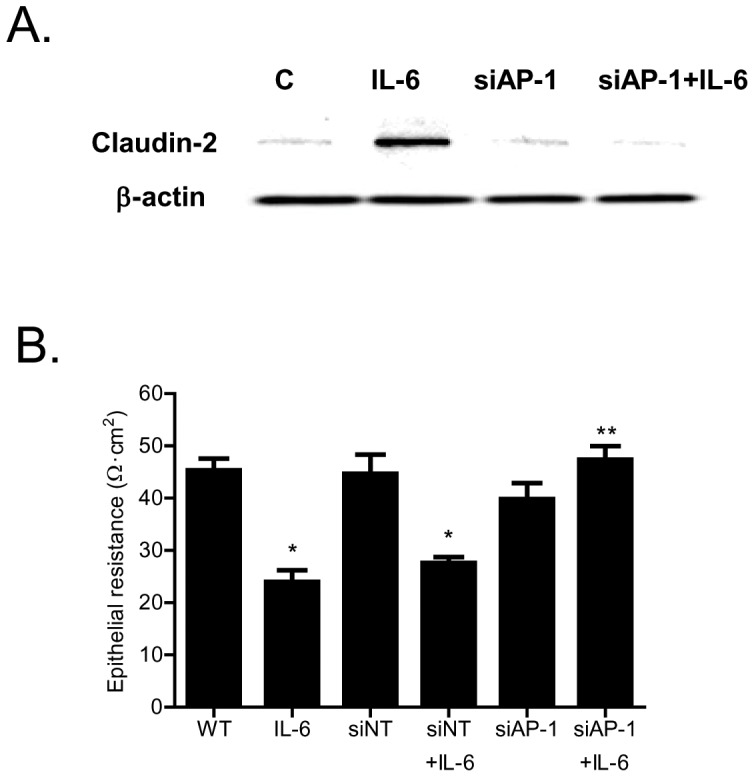
Defective intestinal epithelial tight junction (TJ) barrier has been shown to be a pathogenic factor in the development of intestinal inflammation. Interleukin-6 (IL-6) is a pleiotropic, pro-inflammatory cytokine which plays an important role in promoting inflammatory response in the gut and in the systemic circulation. Despite its key role in mediating variety inflammatory response, the effect of IL-6 on intestinal epithelial barrier remains unclear. The purpose of this study was to investigate the effect of IL-6 on intestinal epithelial TJ barrier and to delineate the intracellular mechanisms involved using in-vitro (filter-grown Caco-2 monolayers) and in-vivo model (mouse intestinal perfusion) systems. Our results indicated that IL-6 causes a site-selective increase in Caco-2 intestinal epithelia TJ permeability, causing an increase in flux of small-sized molecules having molecular radius <4 Å. The size-selective increase in Caco-2 TJ permeability was regulated by protein-specific increase in claudin-2 expression. The IL-6 increase in TJ permeability required activation of JNK signaling cascade. The JNK pathway activation of AP-1 resulted in AP-1 binding to its binding sequence on the claudin-2 promoter region, leading to promoter activation and subsequent increase in claudin-2 gene transcription and protein synthesis and TJ permeability. Our in-vivo mouse perfusion showed that IL-6 modulation of mouse intestinal permeability was also mediated by AP-1 dependent increase in claudin-2 expression. In conclusion, our studies show for the first time that the IL-6 modulation of intestinal TJ permeability was regulated by JNK activation of AP-1 and AP-1 activation of claudin-2 gene.
Conflict of interest statement
Figures
References
-
- Ma TY, Anderson JM (2012) Tight junctions and the intestinal barrier. In: Physiology of the Gastrointestinal Tract,. Burlington, MA: Elsevier Academic Press.
-
- Turner JR (2009) Intestinal mucosal barrier function in health and disease. Nat Rev Immunol 9: 799–809. - PubMed
-
- Piontek J, Winkler L, Wolburg H, Muller SL, Zuleger N, et al. (2008) Formation of tight junction: determinants of homophilic interaction between classic claudins. FASEB J 22: 146–158. - PubMed
-
- Hollander D (1999) Intestinal permeability, leaky gut, and intestinal disorders. Curr Gastroenterol Rep 1: 410–416. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
