

Claudins reign: The claudin/EMP/PMP22/γ channel protein family in C. elegans
- PMID: 24665403
- PMCID: PMC3879130
- DOI: 10.4161/tisb.25502
Claudins reign: The claudin/EMP/PMP22/γ channel protein family in C. elegans
Abstract
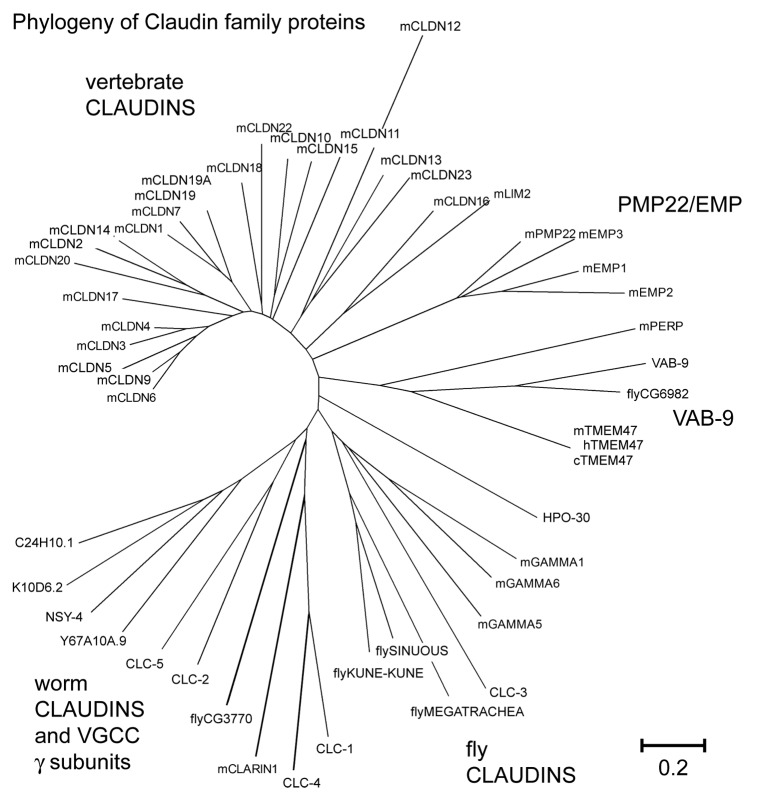
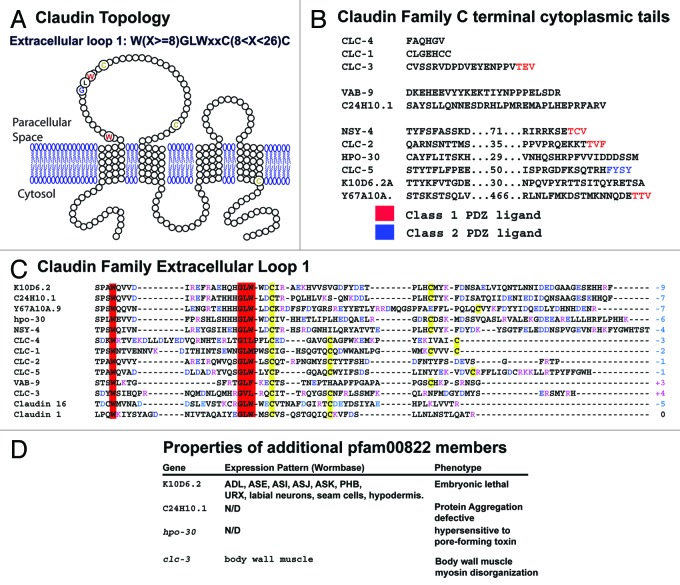
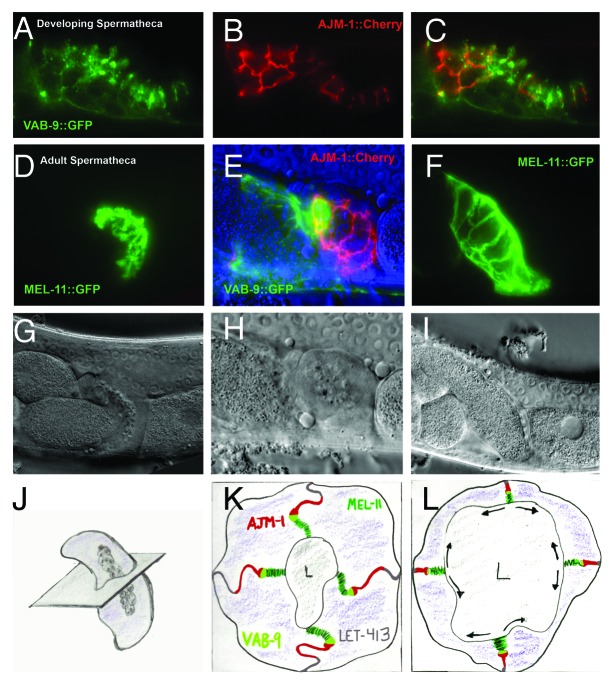
The claudin family of integral membrane proteins was identified as the major protein component of the tight junctions in all vertebrates. Since their identification, claudins, and their associated pfam00822 superfamily of proteins have been implicated in a wide variety of cellular processes. Claudin homologs have been identified in invertebrates as well, including Drosophila and C. elegans. Recent studies demonstrate that the C. elegans claudins, clc-1-clc- 5, and similar proteins in the greater PMP22/EMP/claudin/voltage-gated calcium channel γ subunit family, including nsy-4, and vab-9, while highly divergent at a sequence level from each other and from the vertebrate claudins, in many cases play roles similar to those traditionally assigned to their vertebrate homologs. These include regulating cell adhesion and passage of small molecules through the paracellular space, channel activity, protein aggregation, sensitivity to pore-forming toxins, intercellular signaling, cell fate specification and dynamic changes in cell morphology. Study of claudin superfamily proteins in C. elegans should continue to provide clues as to how claudin family protein function has been adapted to perform diverse functions at specialized cell-cell contacts in metazoans.
Keywords: C. elegans; CLC-1; NSY-4; VAB-9; actomyosin; claudin; epithelia; junctions; morphogenesis; neuronal symmetry.
Figures

References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources