

Depletion of white adipose tissue in cancer cachexia syndrome is associated with inflammatory signaling and disrupted circadian regulation
- PMID: 24667661
- PMCID: PMC3965507
- DOI: 10.1371/journal.pone.0092966
Depletion of white adipose tissue in cancer cachexia syndrome is associated with inflammatory signaling and disrupted circadian regulation
Abstract
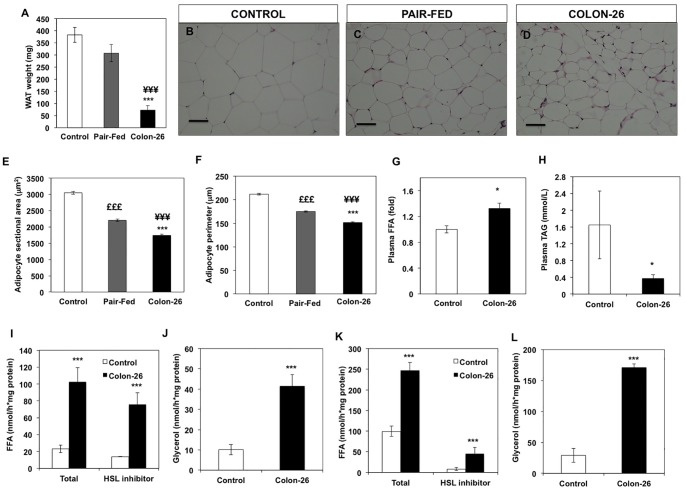
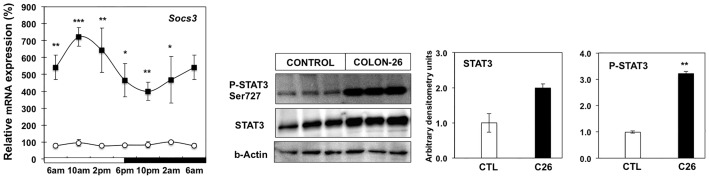
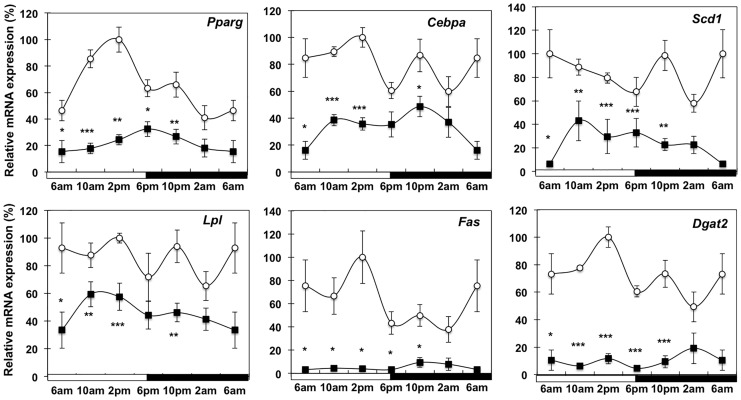
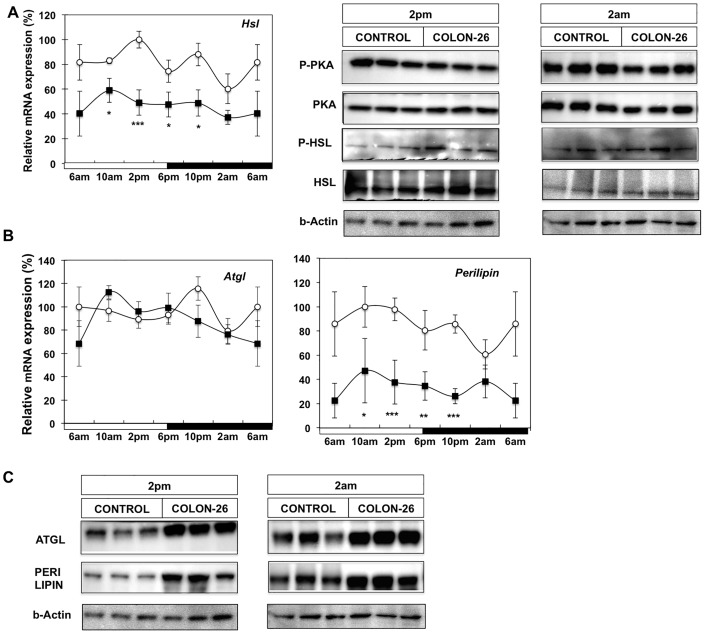
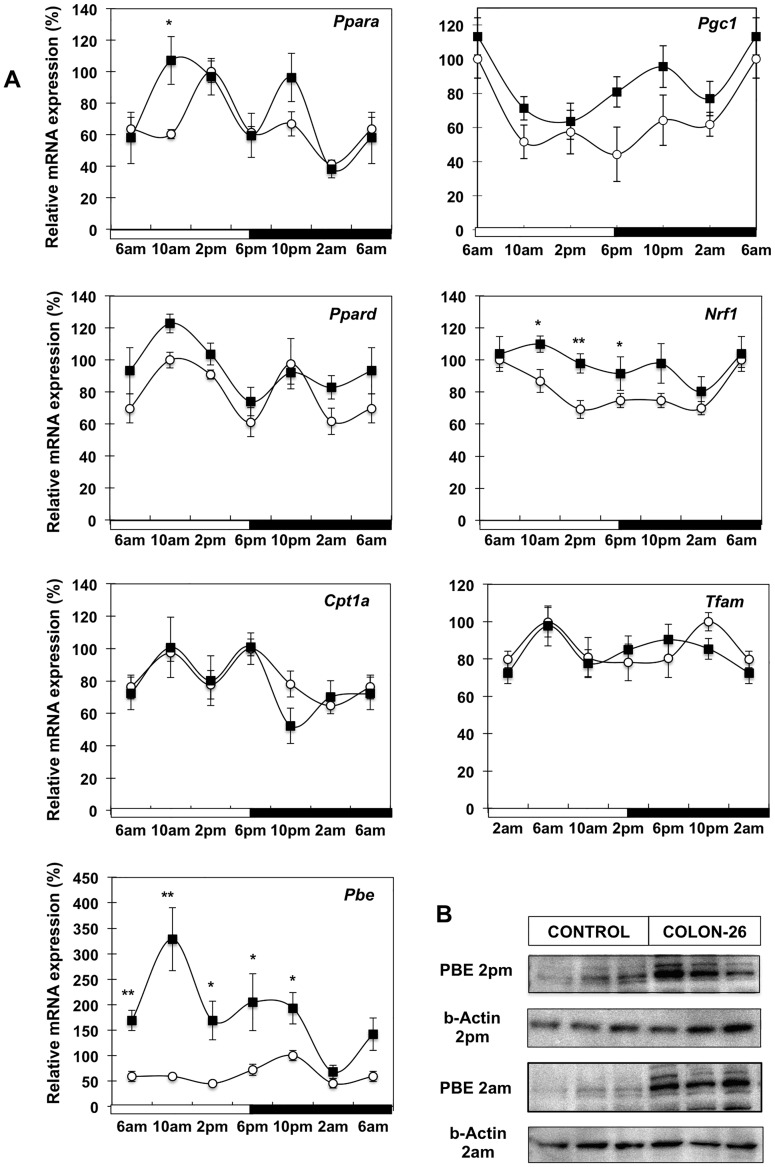
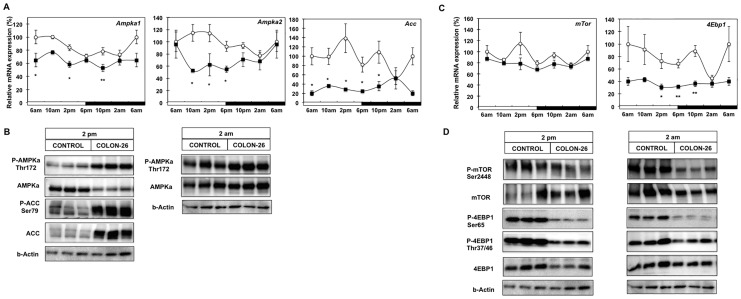
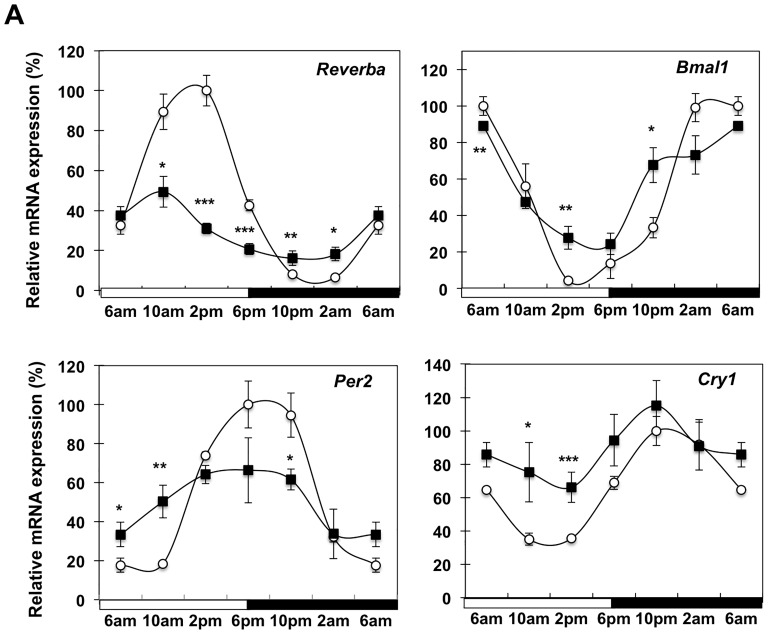
Involuntary weight loss in patients with cancer is the hallmark of cancer cachexia. The etiology of cachexia is multifactorial involving loss of skeletal muscle and adipose tissue associated with high systemic levels of acute phase proteins and inflammatory cytokines. While muscle wasting overtly impacts on cancer patient quality of life, loss of lipid depots represents a sustained energy imbalance. In this study fat depletion was examined in Colon-26 model of cancer cachexia, which is a widely used rodent model of this syndrome. We investigated diurnal expression of circadian rhythm regulators as well as key mediators of energy metabolism and cytokine signaling. Mice bearing the C26 tumour exhibited reduced adipose mass, elevated adipose tissue lipolysis and a 5-fold increase in plasma levels of free fatty acids. These changes were associated with activated IL-6 signaling in WAT through a 3-fold increase in phosphorylated STAT3 and high SOCS3 gene expression levels. In addition perturbations in circadian regulation of lipid metabolism were also observed. Lipid catabolism did not appear to be influenced by the classical PKA pathway activating the lipase HSL. ATGL protein levels were elevated 2-fold in cachectic mice while 4-fold increase phosphorylated ACC and a 2-fold decrease in phosphorylated 4EBP1 was observed indicating that lipid metabolism is modulated by the ATGL & AMPK/mTOR pathways. This study provides evidence for activation of cytokine signaling and concomitant alterations in circadian rhythm and regulators of lipid metabolism in WAT of cachectic animals.
Conflict of interest statement
Figures
References
-
- Palesty JA, Dudrick SJ (2003) What we have learned about cachexia in gastrointestinal cancer. Dig Dis 21: 198–213. - PubMed
-
- Argiles JM, Almendro V, Busquets S, Lopez-Soriano FJ (2004) The pharmacological treatment of cachexia. Curr Drug Targets 5: 265–277. - PubMed
-
- Dewys WD, Begg C, Lavin PT, Bennet PR, Bertino JR (1980) Prognostic effect of weight loss prior to chemotherapy in cancer patients. Am J Med 69: 491–497. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
