

In vivo time-resolved microtomography reveals the mechanics of the blowfly flight motor
- PMID: 24667677
- PMCID: PMC3965381
- DOI: 10.1371/journal.pbio.1001823
In vivo time-resolved microtomography reveals the mechanics of the blowfly flight motor
Abstract
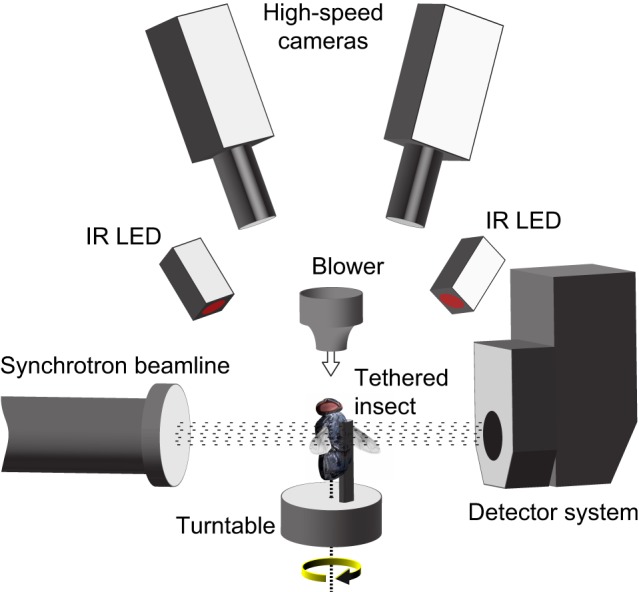
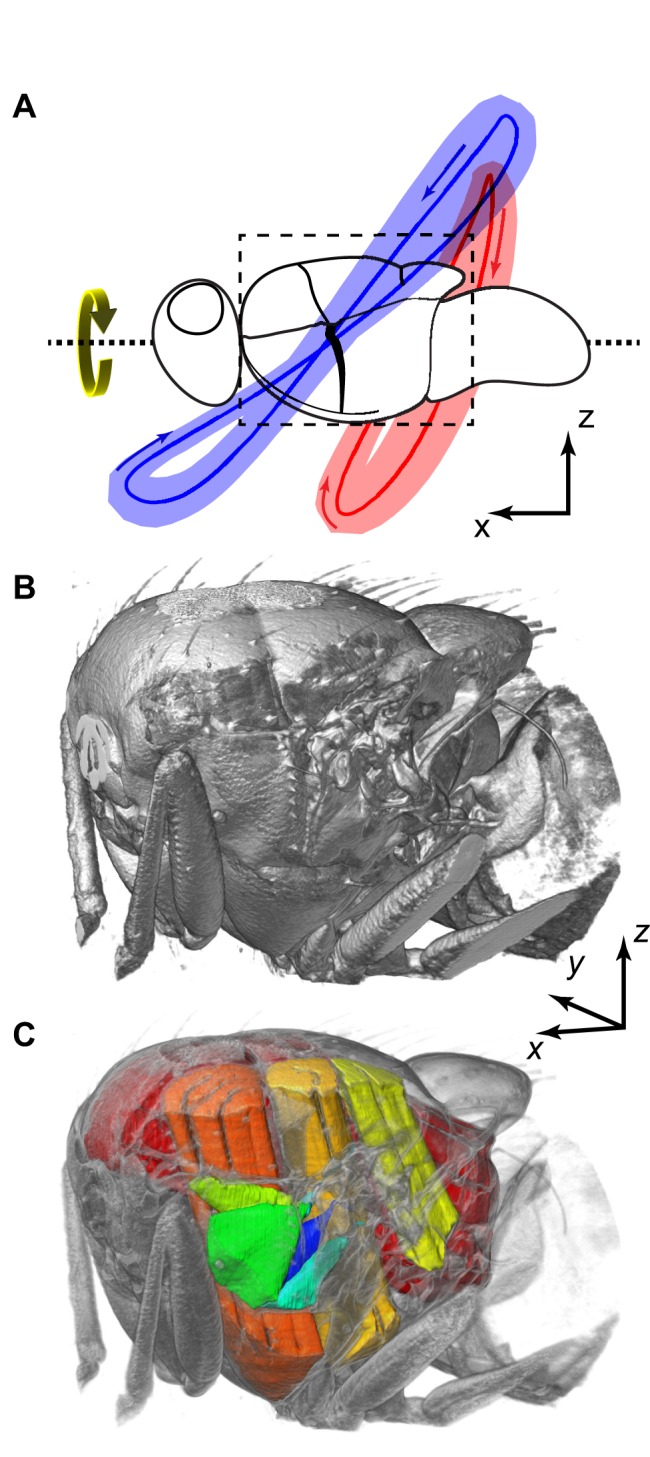
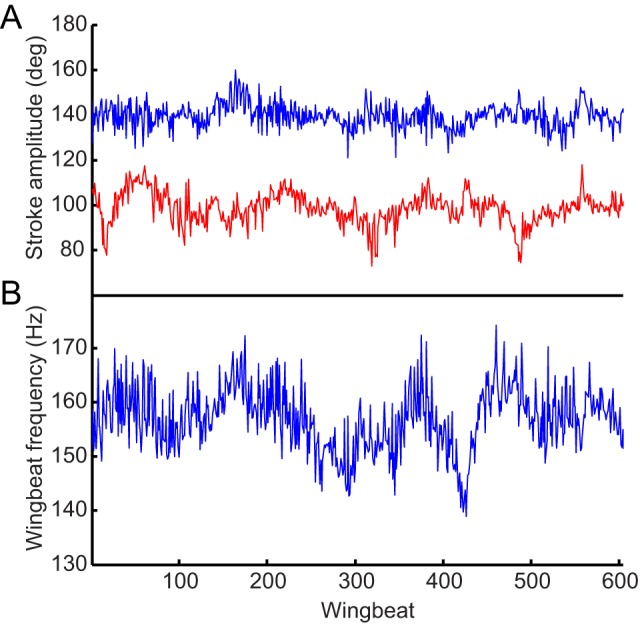
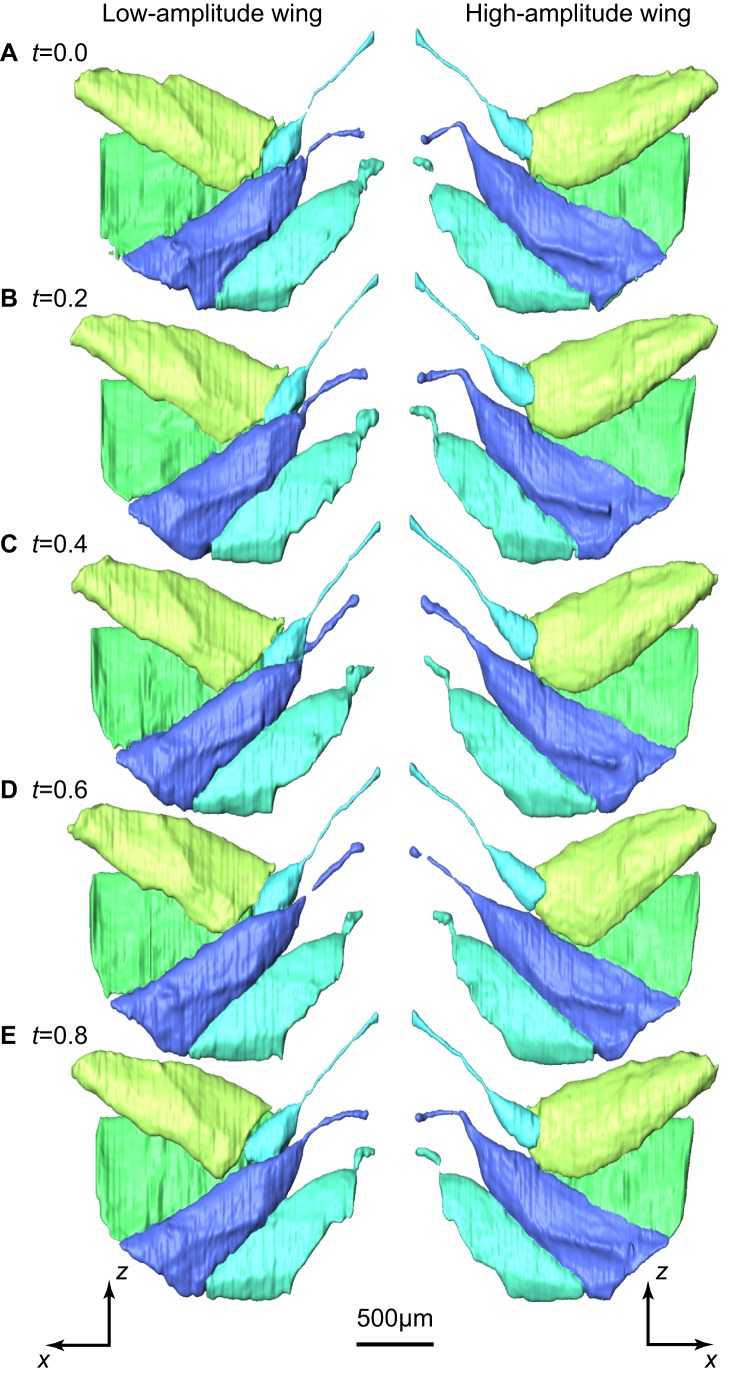
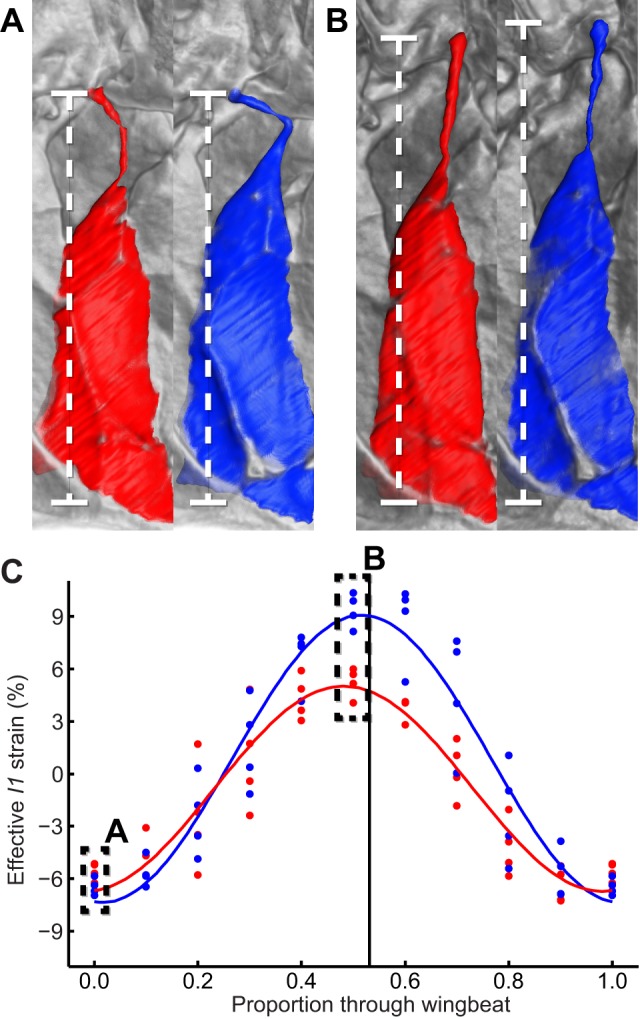
Dipteran flies are amongst the smallest and most agile of flying animals. Their wings are driven indirectly by large power muscles, which cause cyclical deformations of the thorax that are amplified through the intricate wing hinge. Asymmetric flight manoeuvres are controlled by 13 pairs of steering muscles acting directly on the wing articulations. Collectively the steering muscles account for <3% of total flight muscle mass, raising the question of how they can modulate the vastly greater output of the power muscles during manoeuvres. Here we present the results of a synchrotron-based study performing micrometre-resolution, time-resolved microtomography on the 145 Hz wingbeat of blowflies. These data represent the first four-dimensional visualizations of an organism's internal movements on sub-millisecond and micrometre scales. This technique allows us to visualize and measure the three-dimensional movements of five of the largest steering muscles, and to place these in the context of the deforming thoracic mechanism that the muscles actuate. Our visualizations show that the steering muscles operate through a diverse range of nonlinear mechanisms, revealing several unexpected features that could not have been identified using any other technique. The tendons of some steering muscles buckle on every wingbeat to accommodate high amplitude movements of the wing hinge. Other steering muscles absorb kinetic energy from an oscillating control linkage, which rotates at low wingbeat amplitude but translates at high wingbeat amplitude. Kinetic energy is distributed differently in these two modes of oscillation, which may play a role in asymmetric power management during flight control. Structural flexibility is known to be important to the aerodynamic efficiency of insect wings, and to the function of their indirect power muscles. We show that it is integral also to the operation of the steering muscles, and so to the functional flexibility of the insect flight motor.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
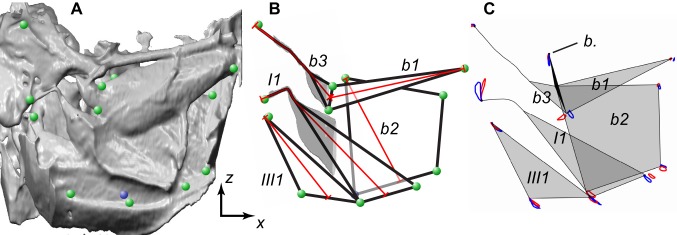
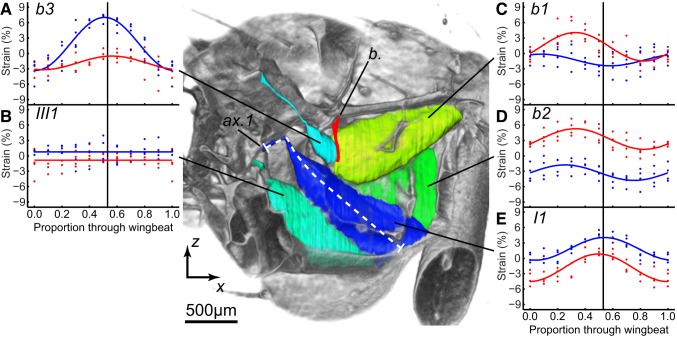
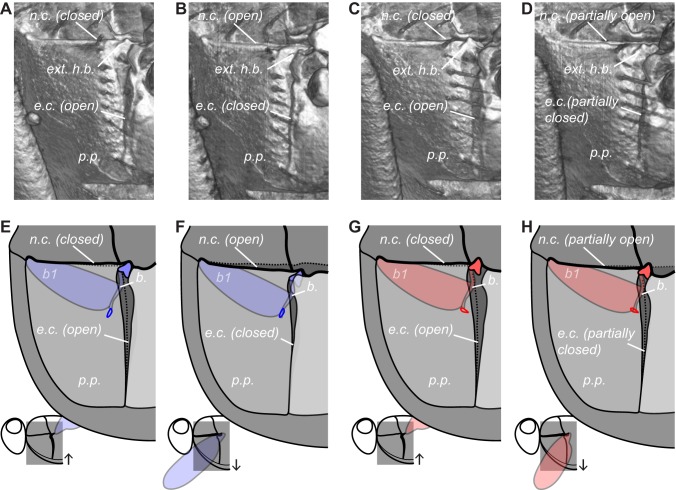
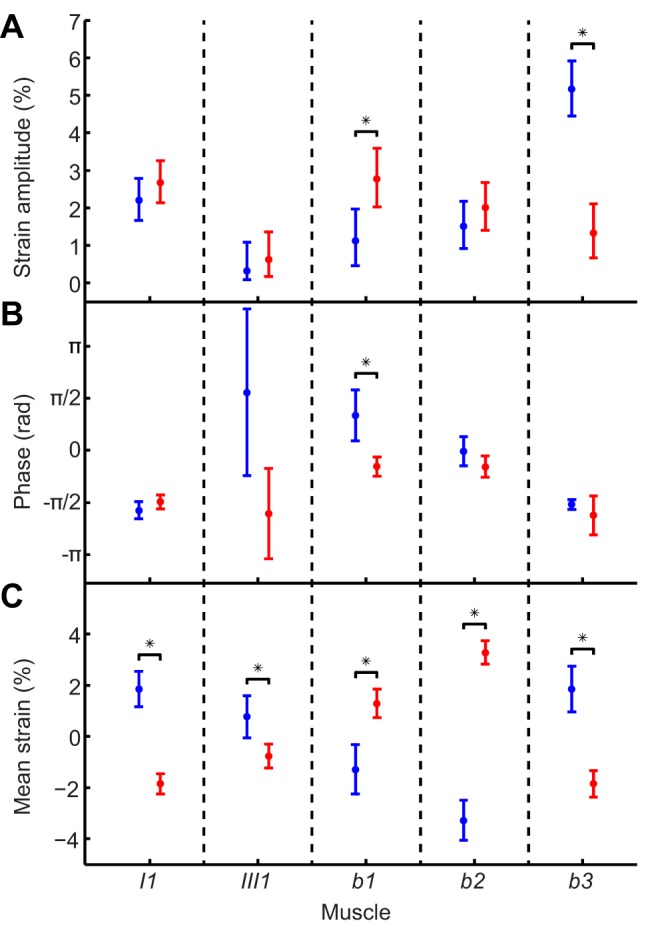
Comment in
-
How insect flight steering muscles work.PLoS Biol. 2014 Mar 25;12(3):e1001822. doi: 10.1371/journal.pbio.1001822. eCollection 2014 Mar. PLoS Biol. 2014. PMID: 24667632 Free PMC article.
References
-
- Pringle JWS (1957) Insect flight. Cambridge: Cambridge University Press.
-
- Dickinson MH, Tu MS (1997) The function of dipteran flight muscle. Comp Biochem Physiol A 116: 223–238.
-
- Heide G (1983) Neural mechanisms of flight control in Diptera. Nachtigall W, editor. BIONA Report 2: Insect flight II. Stuttgart: Fischer. pp. 35–52.
-
- Smyth T (1966) Yurkiewi.Wj (1966) Visual reflex control of indirect flight muscles in the sheep blowfly. Comp Biochem Physiol 17: 1175–1180.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
