

The pathogen Candida albicans hijacks pyroptosis for escape from macrophages
- PMID: 24667705
- PMCID: PMC3977349
- DOI: 10.1128/mBio.00003-14
The pathogen Candida albicans hijacks pyroptosis for escape from macrophages
Abstract
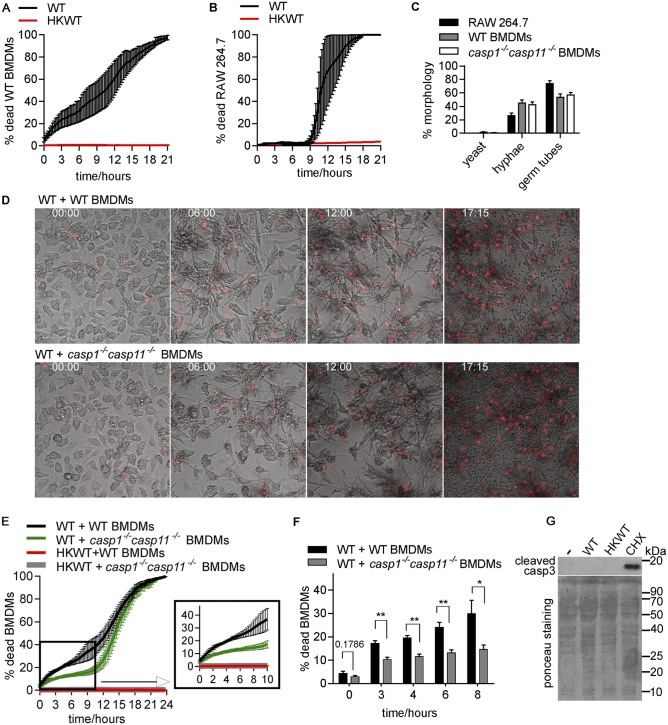
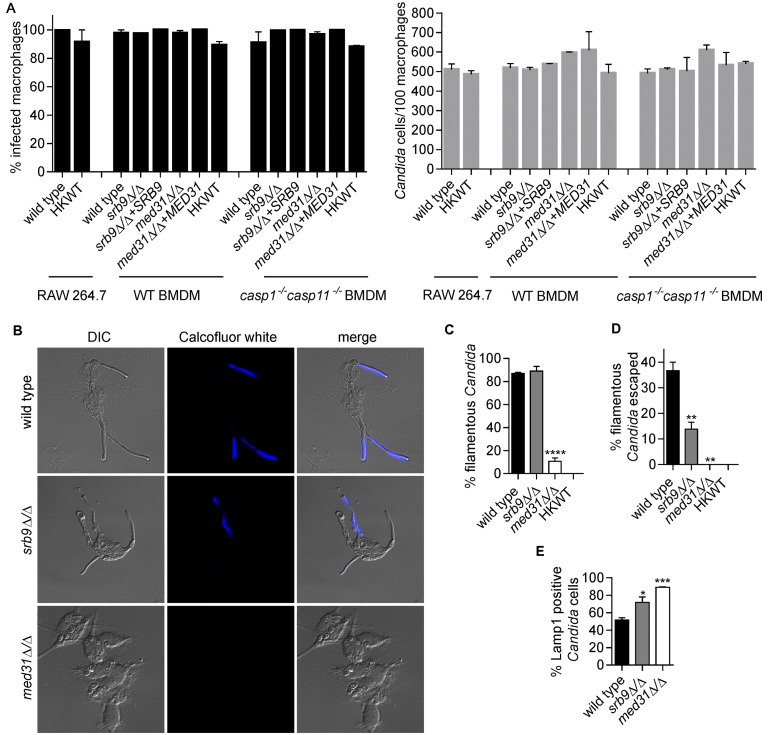
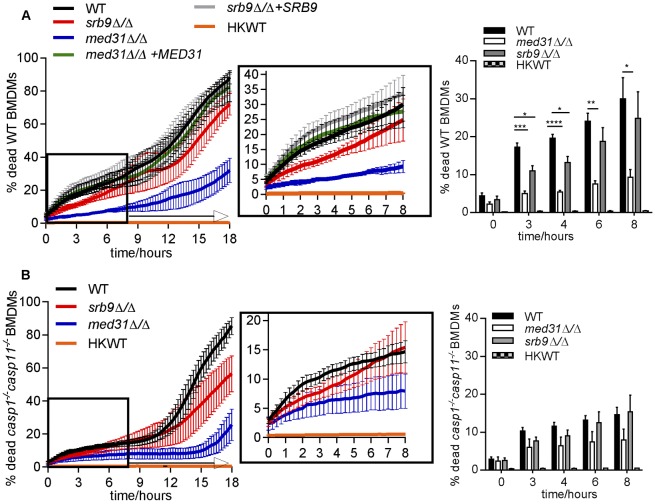
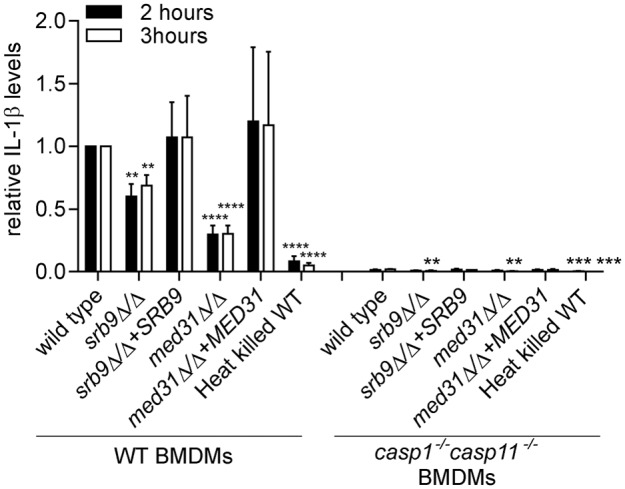
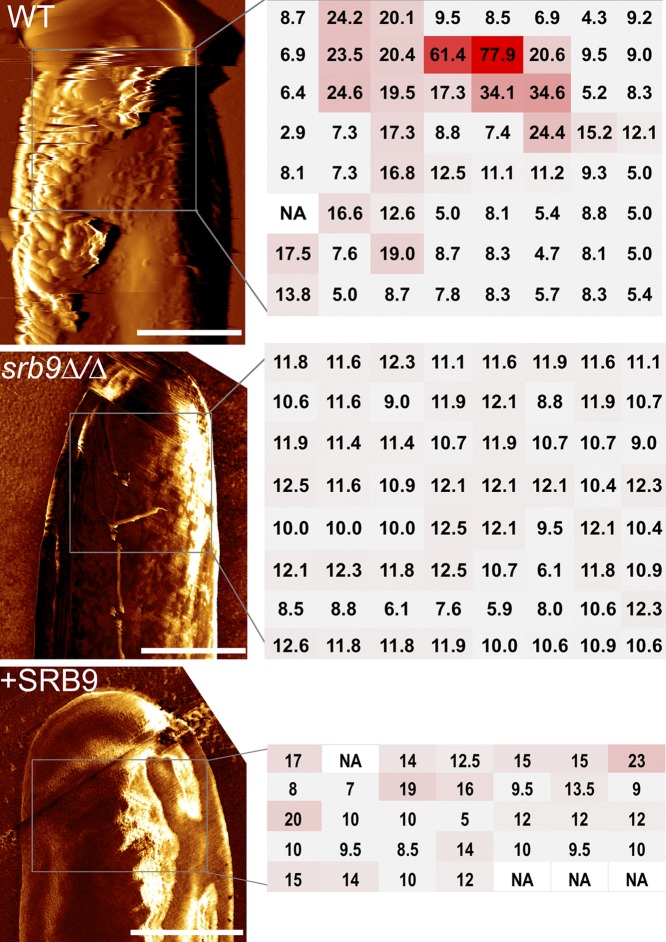
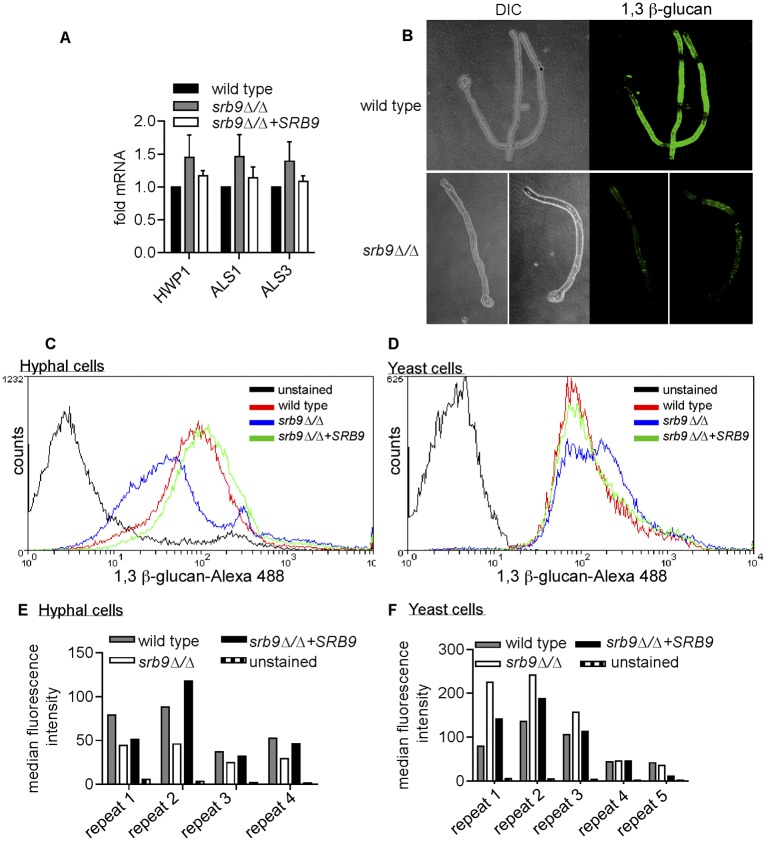
The fungal pathogen Candida albicans causes macrophage death and escapes, but the molecular mechanisms remained unknown. Here we used live-cell imaging to monitor the interaction of C. albicans with macrophages and show that C. albicans kills macrophages in two temporally and mechanistically distinct phases. Early upon phagocytosis, C. albicans triggers pyroptosis, a proinflammatory macrophage death. Pyroptosis is controlled by the developmental yeast-to-hypha transition of Candida. When pyroptosis is inactivated, wild-type C. albicans hyphae cause significantly less macrophage killing for up to 8 h postphagocytosis. After the first 8 h, a second macrophage-killing phase is initiated. This second phase depends on robust hyphal formation but is mechanistically distinct from pyroptosis. The transcriptional regulator Mediator is necessary for morphogenesis of C. albicans in macrophages and the establishment of the wild-type surface architecture of hyphae that together mediate activation of macrophage cell death. Our data suggest that the defects of the Mediator mutants in causing macrophage death are caused, at least in part, by reduced activation of pyroptosis. A Mediator mutant that forms hyphae of apparently wild-type morphology but is defective in triggering early macrophage death shows a breakdown of cell surface architecture and reduced exposed 1,3 β-glucan in hyphae. Our report shows how Candida uses host and pathogen pathways for macrophage killing. The current model of mechanical piercing of macrophages by C. albicans hyphae should be revised to include activation of pyroptosis by hyphae as an important mechanism mediating macrophage cell death upon C. albicans infection. IMPORTANCE Upon phagocytosis by macrophages, Candida albicans can transition to the hyphal form, which causes macrophage death and enables fungal escape. The current model is that the highly polarized growth of hyphae results in macrophage piercing. This model is challenged by recent reports of C. albicans mutants that form hyphae of wild-type morphology but are defective in killing macrophages. We show that C. albicans causes macrophage cell death by at least two mechanisms. Phase 1 killing (first 6 to 8 h) depends on the activation of the pyroptotic programmed host cell death by fungal hyphae. Phase 2 (up to 24 h) is rapid and depends on robust hyphal formation but is independent of pyroptosis. Our data provide a new model for how the interplay between fungal morphogenesis and activation of a host cell death pathway mediates macrophage killing by C. albicans hyphae.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
