

A mouse model for Betacoronavirus subgroup 2c using a bat coronavirus strain HKU5 variant
- PMID: 24667706
- PMCID: PMC3977350
- DOI: 10.1128/mBio.00047-14
A mouse model for Betacoronavirus subgroup 2c using a bat coronavirus strain HKU5 variant
Abstract
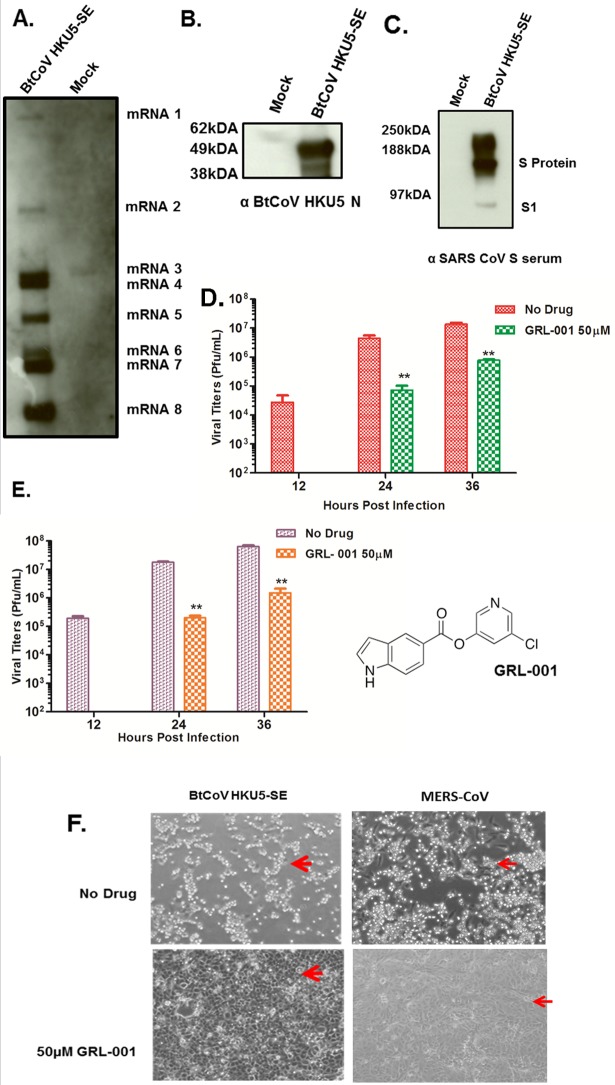
Cross-species transmission of zoonotic coronaviruses (CoVs) can result in pandemic disease outbreaks. Middle East respiratory syndrome CoV (MERS-CoV), identified in 2012, has caused 182 cases to date, with ~43% mortality, and no small animal model has been reported. MERS-CoV and Pipistrellus bat coronavirus (BtCoV) strain HKU5 of Betacoronavirus (β-CoV) subgroup 2c share >65% identity at the amino acid level in several regions, including nonstructural protein 5 (nsp5) and the nucleocapsid (N) protein, which are significant drug and vaccine targets. BtCoV HKU5 has been described in silico but has not been shown to replicate in culture, thus hampering drug and vaccine studies against subgroup 2c β-CoVs. We report the synthetic reconstruction and testing of BtCoV HKU5 containing the severe acute respiratory syndrome (SARS)-CoV spike (S) glycoprotein ectodomain (BtCoV HKU5-SE). This virus replicates efficiently in cell culture and in young and aged mice, where the virus targets airway and alveolar epithelial cells. Unlike some subgroup 2b SARS-CoV vaccines that elicit a strong eosinophilia following challenge, we demonstrate that BtCoV HKU5 and MERS-CoV N-expressing Venezuelan equine encephalitis virus replicon particle (VRP) vaccines do not cause extensive eosinophilia following BtCoV HKU5-SE challenge. Passage of BtCoV HKU5-SE in young mice resulted in enhanced virulence, causing 20% weight loss, diffuse alveolar damage, and hyaline membrane formation in aged mice. Passaged virus was characterized by mutations in the nsp13, nsp14, open reading frame 5 (ORF5) and M genes. Finally, we identified an inhibitor active against the nsp5 proteases of subgroup 2c β-CoVs. Synthetic-genome platforms capable of reconstituting emerging zoonotic viral pathogens or their phylogenetic relatives provide new strategies for identifying broad-based therapeutics, evaluating vaccine outcomes, and studying viral pathogenesis. IMPORTANCE The 2012 outbreak of MERS-CoV raises the specter of another global epidemic, similar to the 2003 SARS-CoV epidemic. MERS-CoV is related to BtCoV HKU5 in target regions that are essential for drug and vaccine testing. Because no small animal model exists to evaluate MERS-CoV pathogenesis or to test vaccines, we constructed a recombinant BtCoV HKU5 that expressed a region of the SARS-CoV spike (S) glycoprotein, thereby allowing the recombinant virus to grow in cell culture and in mice. We show that this recombinant virus targets airway epithelial cells and causes disease in aged mice. We use this platform to (i) identify a broad-spectrum antiviral that can potentially inhibit viruses closely related to MERS-CoV, (ii) demonstrate the absence of increased eosinophilic immune pathology for MERS-CoV N protein-based vaccines, and (iii) mouse adapt this virus to identify viral genetic determinants of cross-species transmission and virulence. This study holds significance as a strategy to control newly emerging viruses.
Figures






References
-
- Pyrc K, Sims AC, Dijkman R, Jebbink M, Long C, Deming D, Donaldson E, Vabret A, Baric R, van der Hoek L, Pickles R. 2010. Culturing the unculturable: human coronavirus HKU1 infects, replicates, and produces progeny virions in human ciliated airway epithelial cell cultures. J. Virol. 84:11255–11263. 10.1128/JVI.00947-10 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
