

Comparison of widely used Listeria monocytogenes strains EGD, 10403S, and EGD-e highlights genomic variations underlying differences in pathogenicity
- PMID: 24667708
- PMCID: PMC3977354
- DOI: 10.1128/mBio.00969-14
Comparison of widely used Listeria monocytogenes strains EGD, 10403S, and EGD-e highlights genomic variations underlying differences in pathogenicity
Abstract
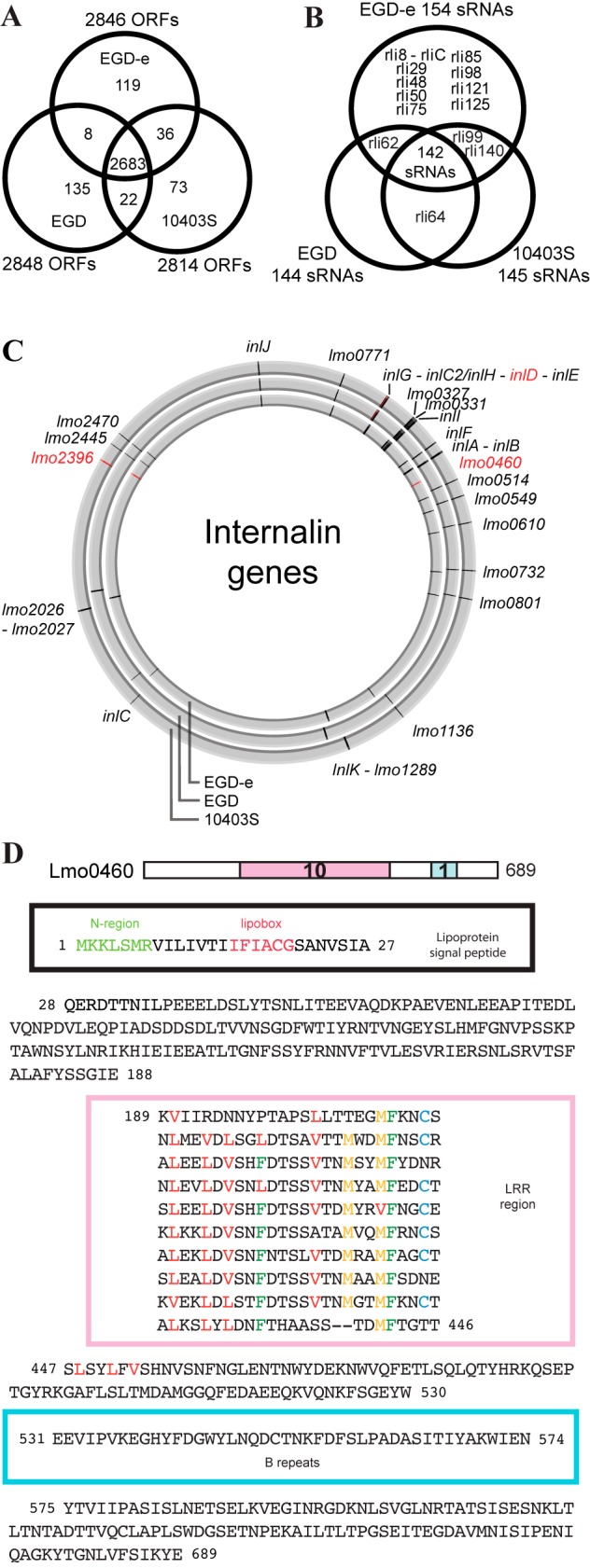
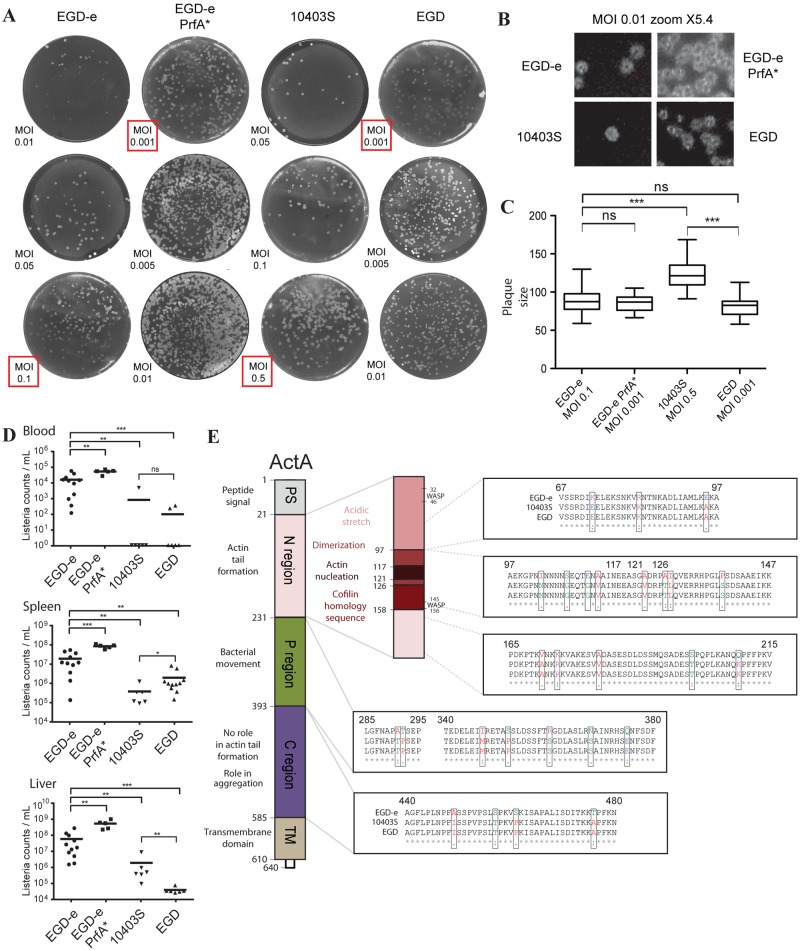
For nearly 3 decades, listeriologists and immunologists have used mainly three strains of the same serovar (1/2a) to analyze the virulence of the bacterial pathogen Listeria monocytogenes. The genomes of two of these strains, EGD-e and 10403S, were released in 2001 and 2008, respectively. Here we report the genome sequence of the third reference strain, EGD, and extensive genomic and phenotypic comparisons of the three strains. Strikingly, EGD-e is genetically highly distinct from EGD (29,016 single nucleotide polymorphisms [SNPs]) and 10403S (30,296 SNPs), and is more related to serovar 1/2c than 1/2a strains. We also found that while EGD and 10403S strains are genetically very close (317 SNPs), EGD has a point mutation in the transcriptional regulator PrfA (PrfA*), leading to constitutive expression of several major virulence genes. We generated an EGD-e PrfA* mutant and showed that EGD behaves like this strain in vitro, with slower growth in broth and higher invasiveness in human cells than those of EGD-e and 10403S. In contrast, bacterial counts in blood, liver, and spleen during infection in mice revealed that EGD and 10403S are less virulent than EGD-e, which is itself less virulent than EGD-e PrfA*. Thus, constitutive expression of PrfA-regulated virulence genes does not appear to provide a significant advantage to the EGD strain during infection in vivo, highlighting the fact that in vitro invasion assays are not sufficient for evaluating the pathogenic potential of L. monocytogenes strains. Together, our results pave the way for deciphering unexplained differences or discrepancies in experiments using different L. monocytogenes strains. IMPORTANCE Over the past 3 decades, Listeria has become a model organism for host-pathogen interactions, leading to critical discoveries in a broad range of fields, including bacterial gene regulation, cell biology, and bacterial pathophysiology. Scientists studying Listeria use primarily three pathogenic strains: EGD, EGD-e, and 10403S. Despite many studies on EGD, it is the only one of the three strains whose genome has not been sequenced. Here we report the sequence of its genome and a series of important genomic and phenotypic differences between the three strains, in particular, a critical mutation in EGD's PrfA, the main regulator of Listeria virulence. Our results show that the three strains display differences which may play an important role in the virulence differences observed between the strains. Our findings will be of critical relevance to listeriologists and immunologists who have used or may use Listeria as a tool to study the pathophysiology of listeriosis and immune responses.
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
