

Mechanisms of perinatal arterial ischemic stroke
- PMID: 24667913
- PMCID: PMC4050239
- DOI: 10.1038/jcbfm.2014.41
Mechanisms of perinatal arterial ischemic stroke
Abstract
The incidence of perinatal stroke is high, similar to that in the elderly, and produces a significant morbidity and severe long-term neurologic and cognitive deficits, including cerebral palsy, epilepsy, neuropsychological impairments, and behavioral disorders. Emerging clinical data and data from experimental models of cerebral ischemia in neonatal rodents have shown that the pathophysiology of perinatal brain damage is multifactorial. These studies have revealed that, far from just being a smaller version of the adult brain, the neonatal brain is unique with a very particular and age-dependent responsiveness to hypoxia-ischemia and focal arterial stroke. In this review, we discuss fundamental clinical aspects of perinatal stroke as well as some of the most recent and relevant findings regarding the susceptibility of specific brain cell populations to injury, the dynamics and the mechanisms of neuronal cell death in injured neonates, the responses of neonatal blood-brain barrier to stroke in relation to systemic and local inflammation, and the long-term effects of stroke on angiogenesis and neurogenesis. Finally, we address translational strategies currently being considered for neonatal stroke as well as treatments that might effectively enhance repair later after injury.
Figures
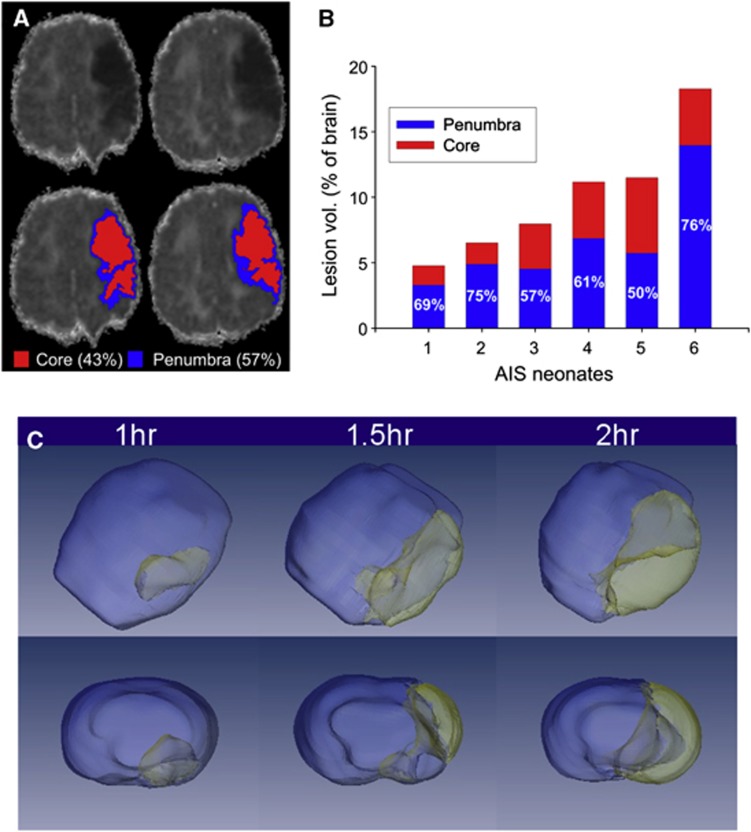
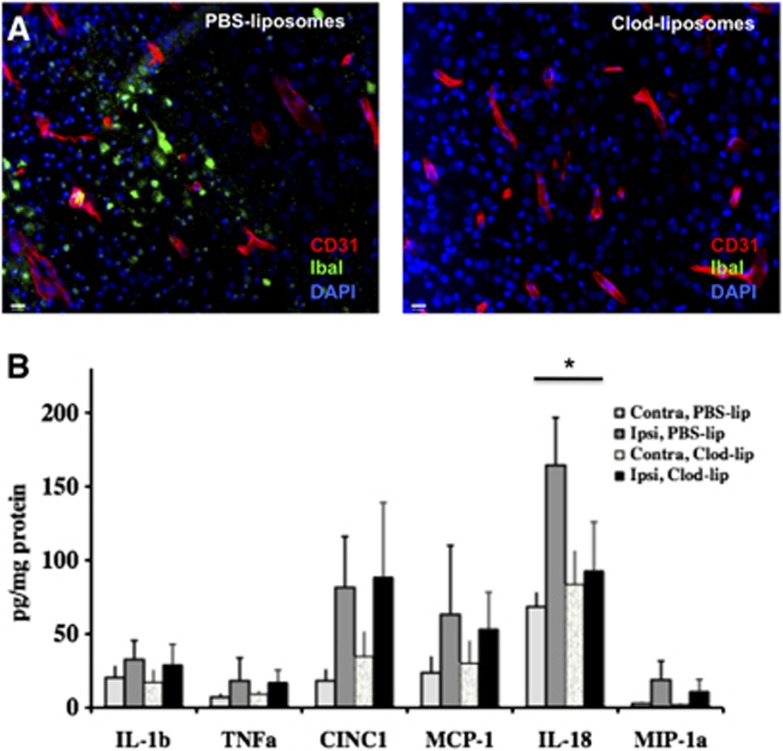
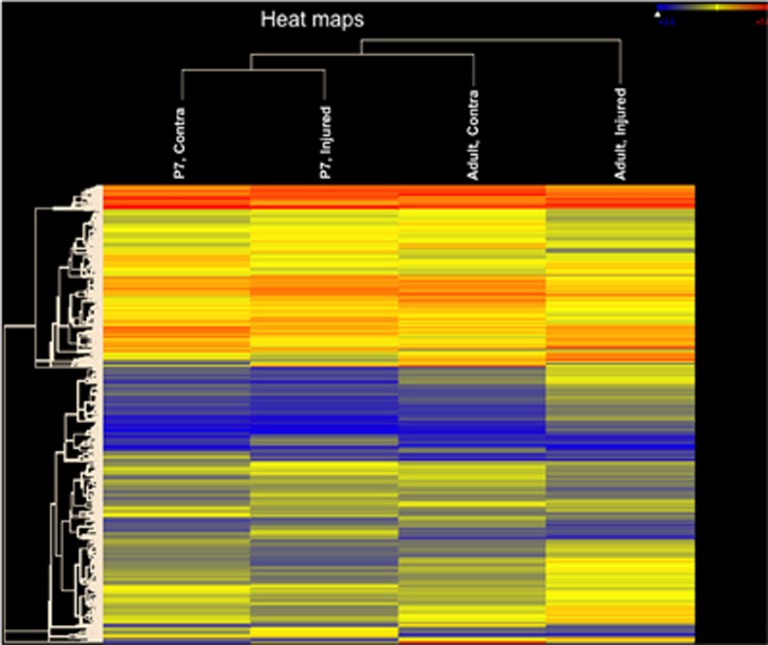
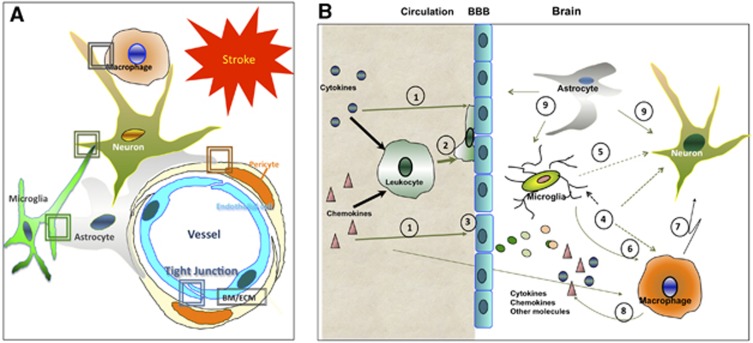
Similar articles
-
Barrier mechanisms in neonatal stroke.Front Neurosci. 2014 Nov 7;8:359. doi: 10.3389/fnins.2014.00359. eCollection 2014. Front Neurosci. 2014. PMID: 25426016 Free PMC article. Review.
-
Systemic Proteasome Inhibition Induces Sustained Post-stroke Neurological Recovery and Neuroprotection via Mechanisms Involving Reversal of Peripheral Immunosuppression and Preservation of Blood-Brain-Barrier Integrity.Mol Neurobiol. 2016 Nov;53(9):6332-6341. doi: 10.1007/s12035-015-9533-3. Epub 2015 Nov 16. Mol Neurobiol. 2016. PMID: 26572637
-
Pathophysiology and neuroprotection of global and focal perinatal brain injury: lessons from animal models.Pediatr Neurol. 2015 Jun;52(6):566-584. doi: 10.1016/j.pediatrneurol.2015.01.016. Epub 2015 Jan 31. Pediatr Neurol. 2015. PMID: 26002050 Free PMC article. Review.
-
Blood-brain barrier, reperfusion injury, and hemorrhagic transformation in acute ischemic stroke.Neurology. 2012 Sep 25;79(13 Suppl 1):S52-7. doi: 10.1212/WNL.0b013e3182697e70. Neurology. 2012. PMID: 23008413 Review. No abstract available.
-
Ischemic stroke in neonatal and adult astrocytes.Mech Ageing Dev. 2019 Oct;183:111147. doi: 10.1016/j.mad.2019.111147. Epub 2019 Sep 4. Mech Ageing Dev. 2019. PMID: 31493435 Review.
Cited by
-
Electroacupuncture on the Scalp over the Motor Cortex Ameliorates Behavioral Deficits Following Neonatal Hypoxia-Ischemia in Rats via the Activation of Neural Stem Cells.Life (Basel). 2020 Oct 14;10(10):240. doi: 10.3390/life10100240. Life (Basel). 2020. PMID: 33066563 Free PMC article.
-
Focal Ischemic Injury to the Early Neonatal Rat Brain Models Cognitive and Motor Deficits with Associated Histopathological Outcomes Relevant to Human Neonatal Brain Injury.Int J Mol Sci. 2021 Apr 29;22(9):4740. doi: 10.3390/ijms22094740. Int J Mol Sci. 2021. PMID: 33947043 Free PMC article.
-
Neuroregeneration and functional recovery after stroke: advancing neural stem cell therapy toward clinical application.Neural Regen Res. 2021 Jan;16(1):80-92. doi: 10.4103/1673-5374.286955. Neural Regen Res. 2021. PMID: 32788451 Free PMC article.
-
Neuroprotection of the Perinatal Brain by Early Information of Cerebral Oxygenation and Perfusion Patterns.Int J Mol Sci. 2021 May 20;22(10):5389. doi: 10.3390/ijms22105389. Int J Mol Sci. 2021. PMID: 34065460 Free PMC article. Review.
-
The Ischemic Immature Brain: Views on Current Experimental Models.Front Cell Neurosci. 2018 Aug 29;12:277. doi: 10.3389/fncel.2018.00277. eCollection 2018. Front Cell Neurosci. 2018. PMID: 30210301 Free PMC article. No abstract available.
References
-
- Nelson KB. Perinatal ischemic stroke. Stroke. 2007;38:742–745. - PubMed
-
- Lynch JK, Nelson KB. Epidemiology of perinatal stroke. Curr Opin Pediatr. 2001;13:499–505. - PubMed
-
- Nelson KB, Lynch JK. Stroke in newborn infants. Lancet Neurol. 2004;3:150–158. - PubMed
-
- Raju TN, Nelson KB, Ferriero D, Lynch JK. Ischemic perinatal stroke: summary of a workshop sponsored by the National Institute of Child Health and Human Development and the National Institute of Neurological Disorders and Stroke. Pediatrics. 2007;120:609–616. - PubMed
-
- Lee J, Croen LA, Lindan C, Nash KB, Yoshida CK, Ferriero DM, et al. Predictors of outcome in perinatal arterial stroke: a population-based study. Ann Neurol. 2005;58:303–308. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
