

Alcohol dehydrogenase III exacerbates liver fibrosis by enhancing stellate cell activation and suppressing natural killer cells in mice
- PMID: 24668648
- PMCID: PMC4867000
- DOI: 10.1002/hep.27137
Alcohol dehydrogenase III exacerbates liver fibrosis by enhancing stellate cell activation and suppressing natural killer cells in mice
Abstract
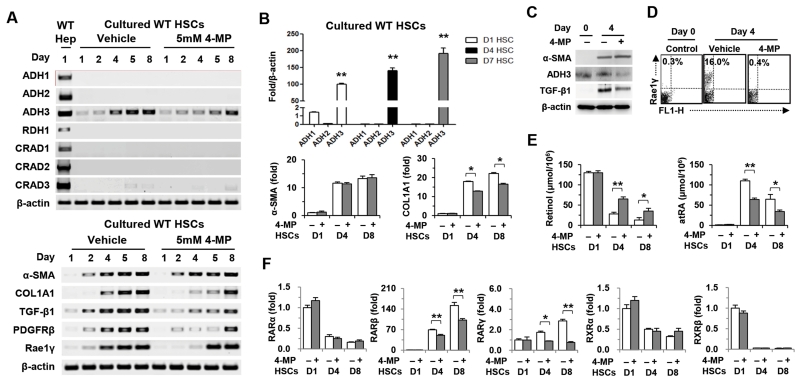
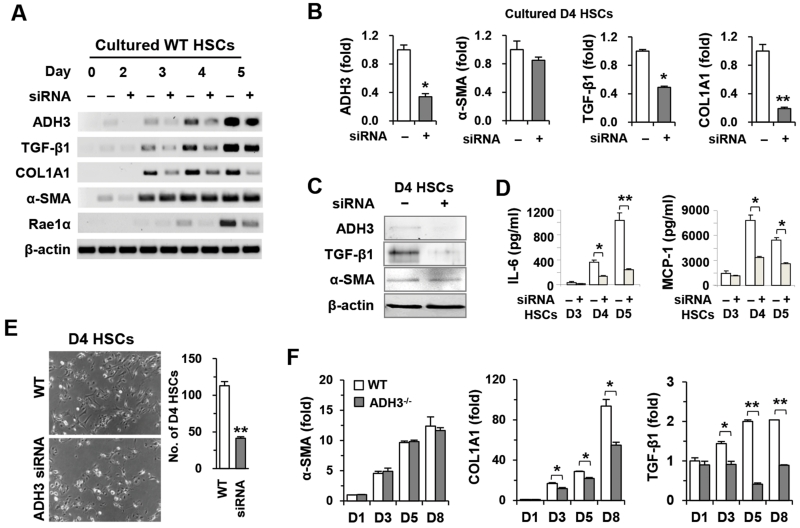
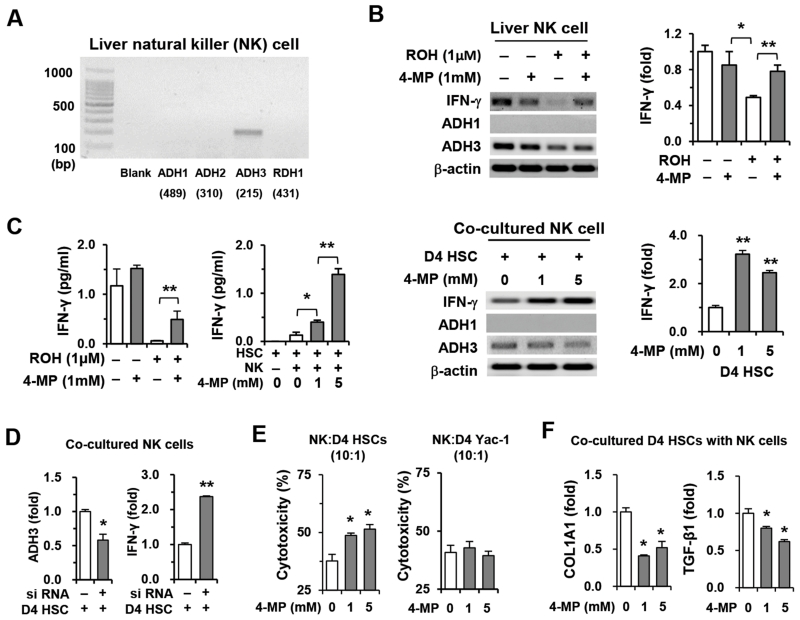
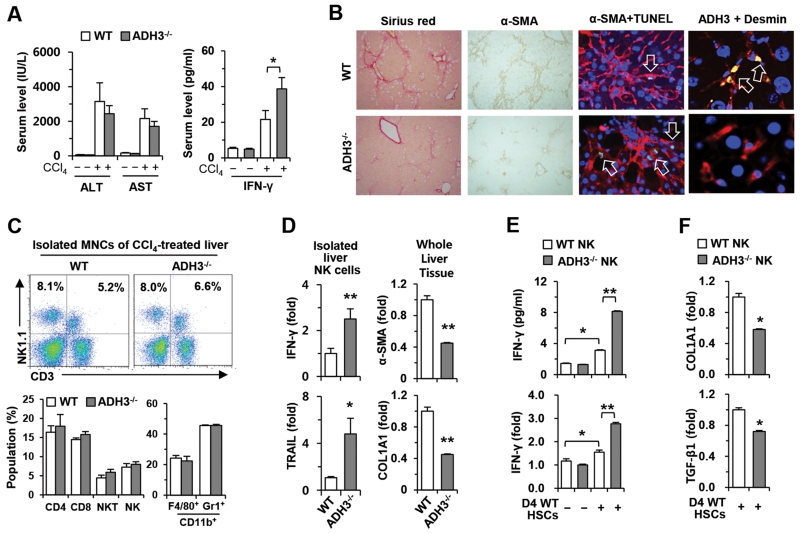
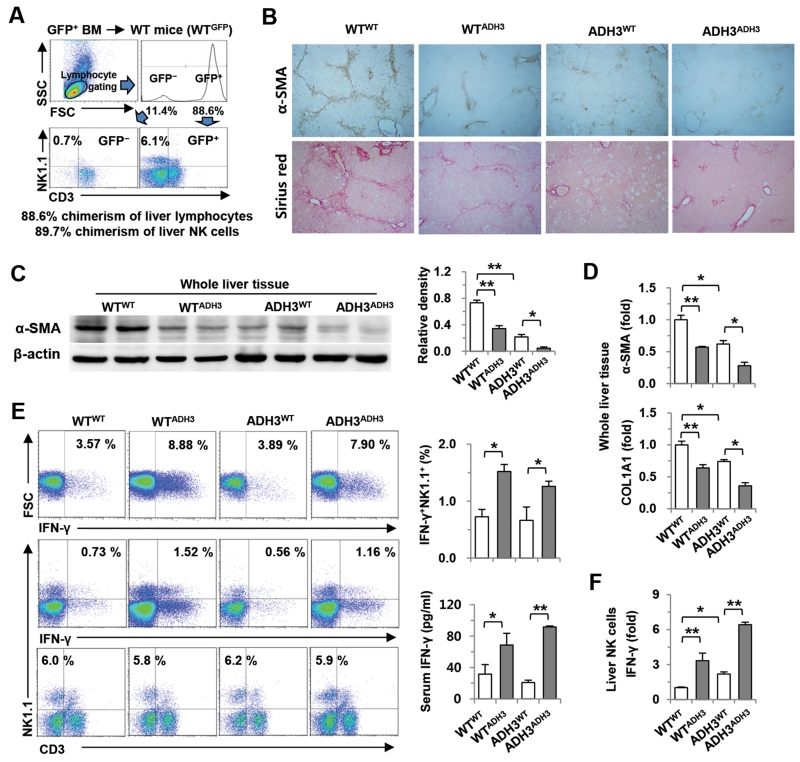
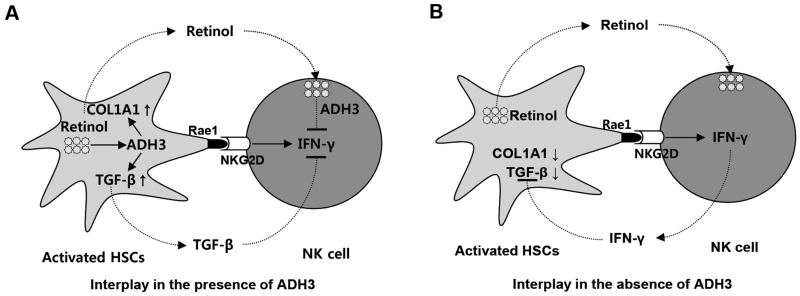
The important roles of retinols and their metabolites have recently been emphasized in the interactions between hepatic stellate cells (HSCs) and natural killer (NK) cells. Nevertheless, the expression and role of retinol metabolizing enzyme in both cell types have yet to be clarified. Thus, we investigated the expression of retinol metabolizing enzyme and its role in liver fibrosis. Among several retinol metabolizing enzymes, only alcohol dehydrogenase (ADH) 3 expression was detected in isolated HSCs and NK cells, whereas hepatocytes express all of them. In vitro treatment with 4-methylpyrazole (4-MP), a broad ADH inhibitor, or depletion of the ADH3 gene down-regulated collagen and transforming growth factor-β1 (TGF-β1) gene expression, but did not affect α-smooth muscle actin gene expression in cultured HSCs. Additionally, in vitro, treatments with retinol suppressed NK cell activities, whereas inhibition of ADH3 enhanced interferon-γ (IFN-γ) production and cytotoxicity of NK cells against HSCs. In vivo, genetic depletion of the ADH3 gene ameliorated bile duct ligation- and carbon tetrachloride-induced liver fibrosis, in which a higher number of apoptotic HSCs and an enhanced activation of NK cells were detected. Freshly isolated HSCs from ADH3-deficient mice showed reduced expression of collagen and TGF-β1, but enhanced expression of IFN-γ was detected in NK cells from these mice compared with those of control mice. Using reciprocal bone marrow transplantation of wild-type and ADH3-deficient mice, we demonstrated that ADH3 deficiency in both HSCs and NK cells contributed to the suppressed liver fibrosis.
Conclusion: ADH3 plays important roles in promoting liver fibrosis by enhancing HSC activation and inhibiting NK cell activity, and could be used as a potential therapeutic target for the treatment of liver fibrosis.
© 2014 by the American Association for the Study of Liver Diseases.
Figures
References
-
- Radaeva S, Wang L, Radaev S, Jeong WI, Park O, Gao B. Retinoic acid signaling sensitizes hepatic stellate cells to NK cell killing via upregulation of NK cell activating ligand RAE1. Am J Physiol Gastrointest Liver Physiol. 2007;293:G809–816. - PubMed
-
- Duester G, Mic FA, Molotkov A. Cytosolic retinoid dehydrogenases govern ubiquitous metabolism of retinol to retinaldehyde followed by tissue-specific metabolism to retinoic acid. Chem Biol Interact. 2003;143-144:201–210. - PubMed
-
- Crabb DW, Matsumoto M, Chang D, You M. Overview of the role of alcohol dehydrogenase and aldehyde dehydrogenase and their variants in the genesis of alcohol-related pathology. Proc Nutr Soc. 2004;63:49–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases