

miRNAs trigger widespread epigenetically activated siRNAs from transposons in Arabidopsis
- PMID: 24670663
- PMCID: PMC4074602
- DOI: 10.1038/nature13069
miRNAs trigger widespread epigenetically activated siRNAs from transposons in Arabidopsis
Abstract
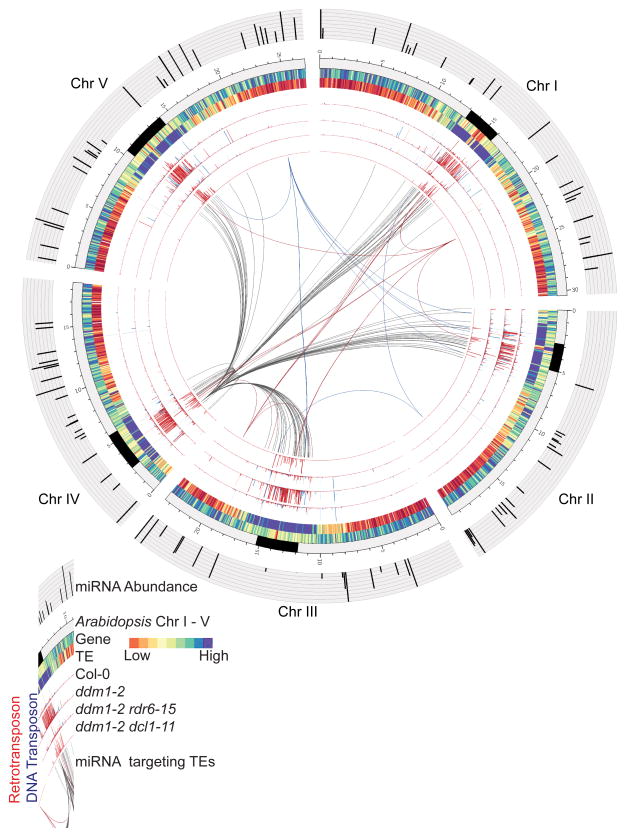
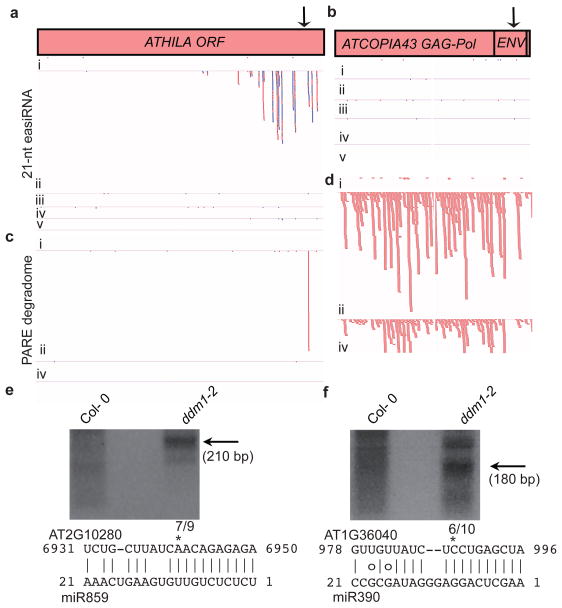
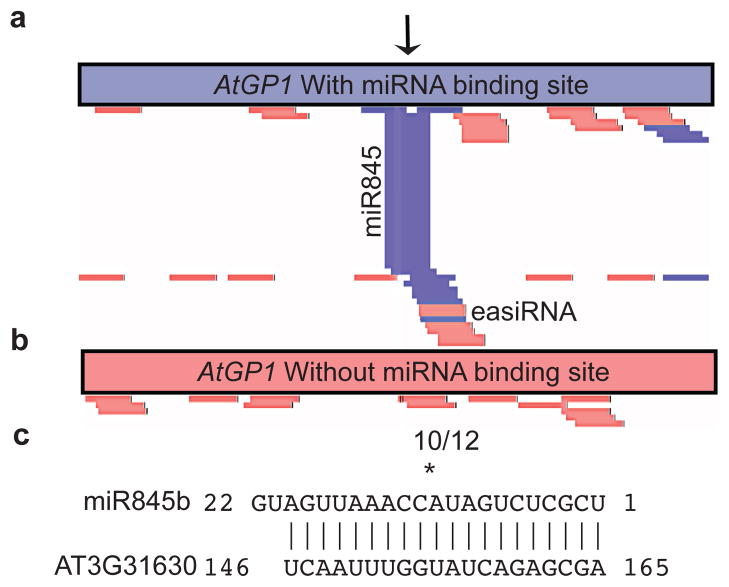
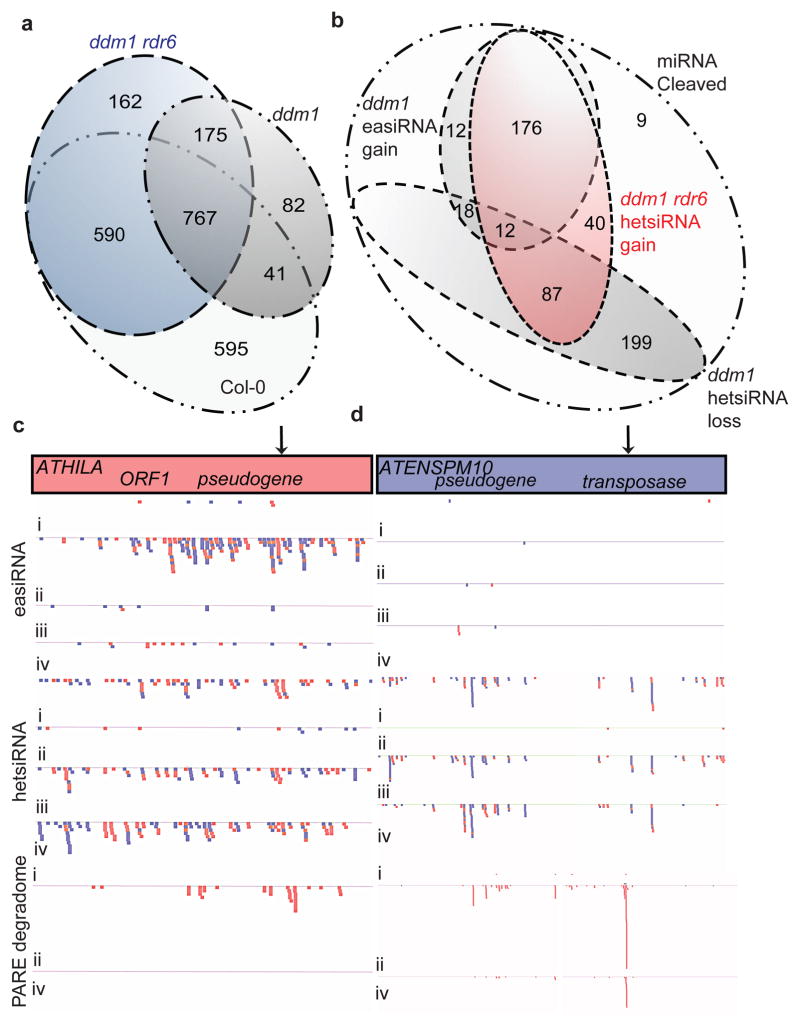
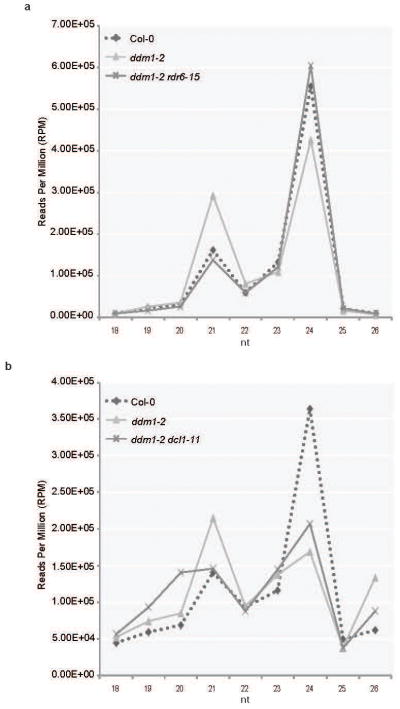
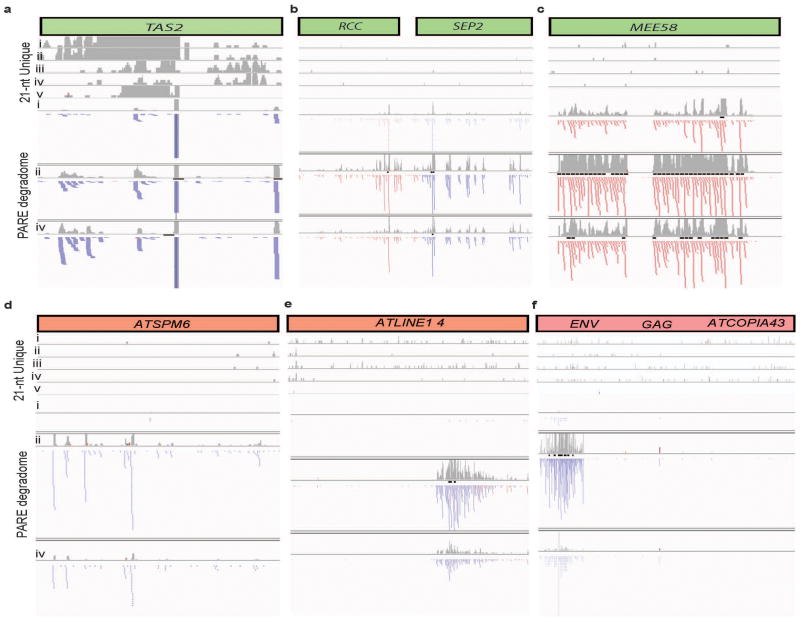
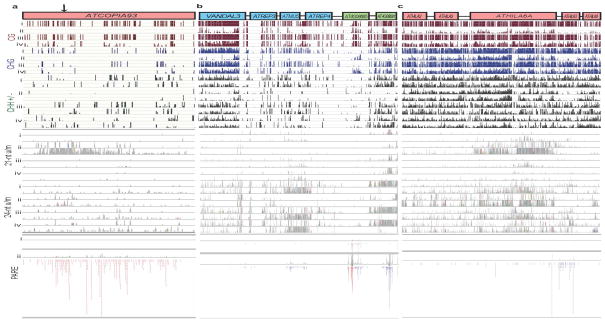
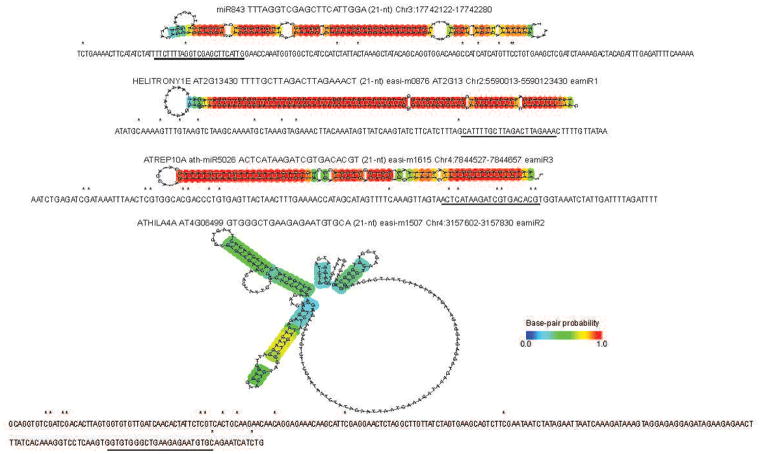
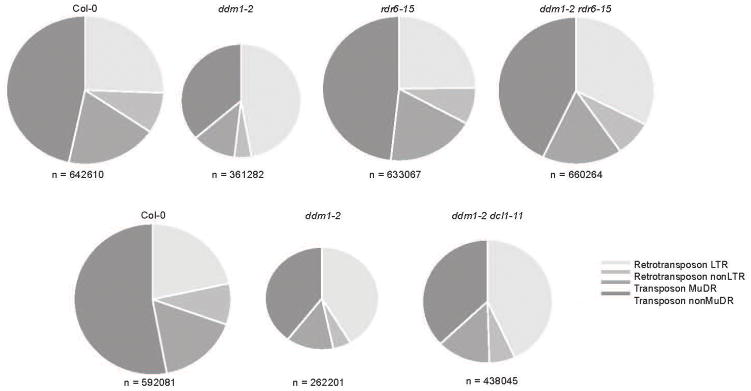
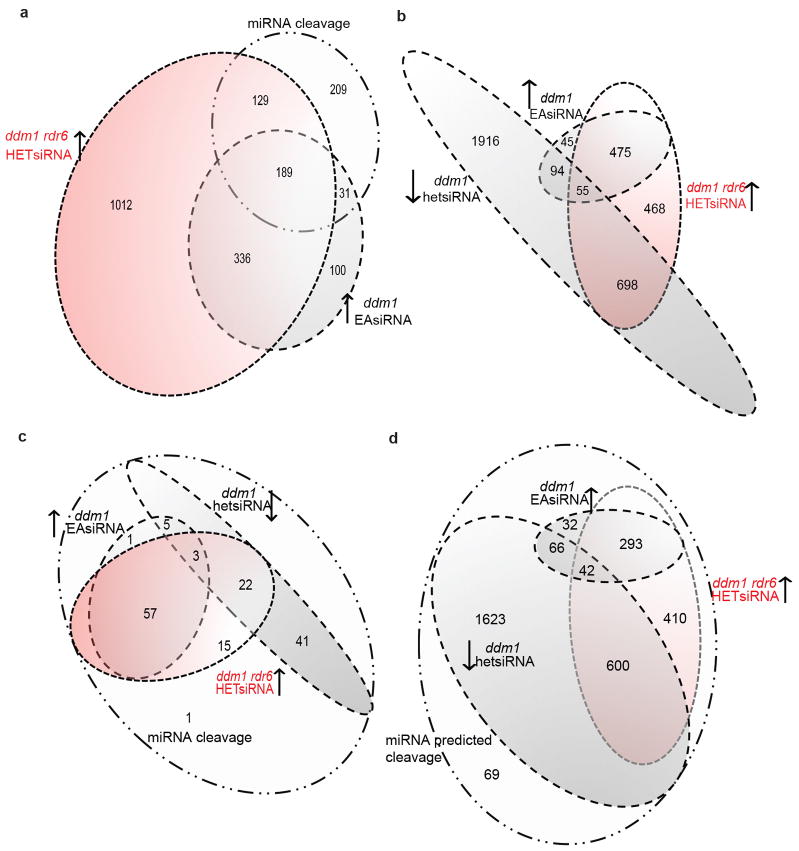
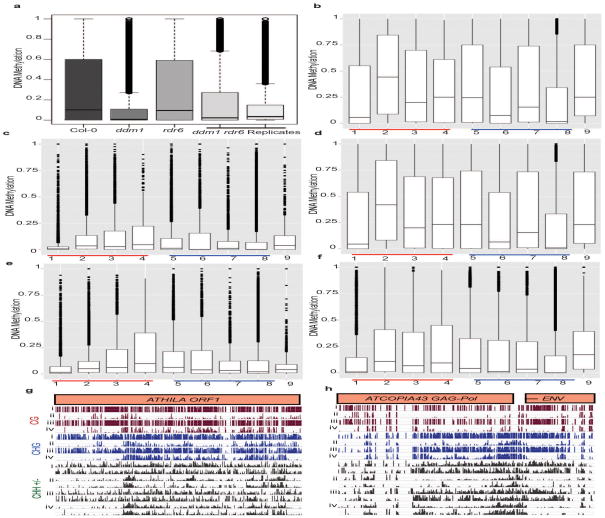
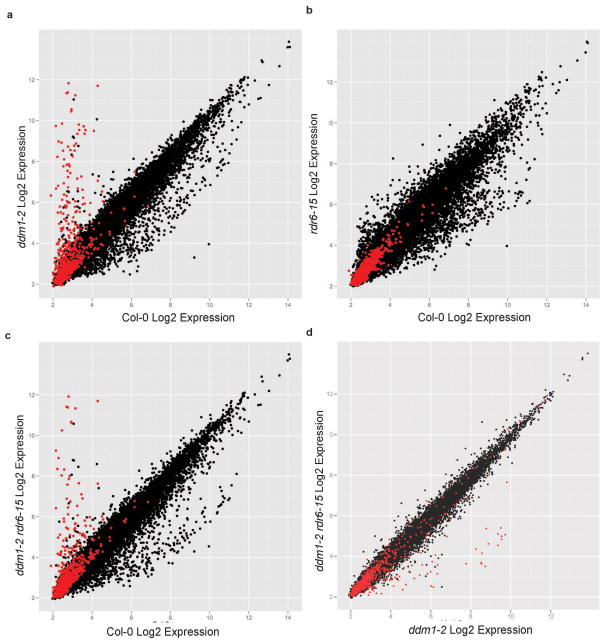
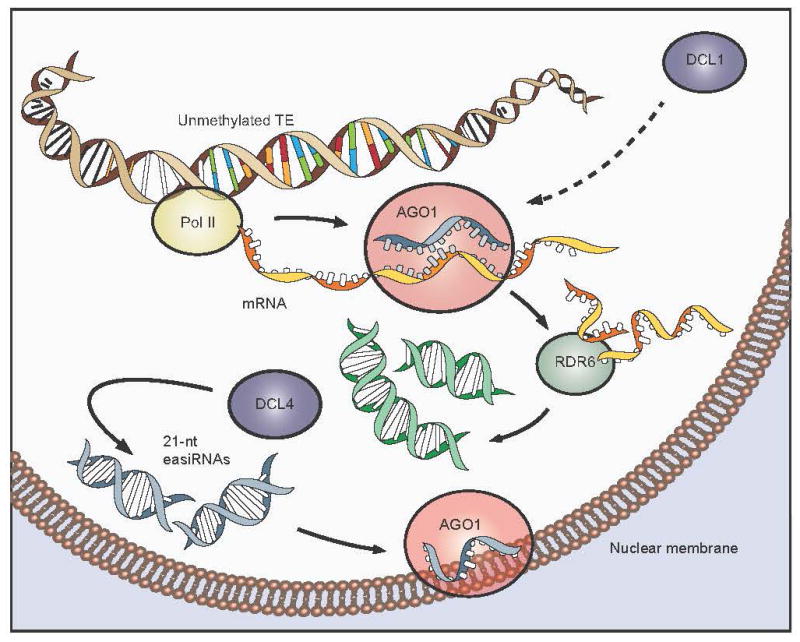
In plants, post-transcriptional gene silencing (PTGS) is mediated by DICER-LIKE 1 (DCL1)-dependent microRNAs (miRNAs), which also trigger 21-nucleotide secondary short interfering RNAs (siRNAs) via RNA-DEPENDENT RNA POLYMERASE 6 (RDR6), DCL4 and ARGONAUTE 1 (AGO1), whereas transcriptional gene silencing (TGS) of transposons is mediated by 24-nucleotide heterochromatic (het)siRNAs, RDR2, DCL3 and AGO4 (ref. 4). Transposons can also give rise to abundant 21-nucleotide 'epigenetically activated' small interfering RNAs (easiRNAs) in DECREASED DNA METHYLATION 1 (ddm1) and DNA METHYLTRANSFERASE 1 (met1) mutants, as well as in the vegetative nucleus of pollen grains and in dedifferentiated plant cell cultures. Here we show that easiRNAs in Arabidopsis thaliana resemble secondary siRNAs, in that thousands of transposon transcripts are specifically targeted by more than 50 miRNAs for cleavage and processing by RDR6. Loss of RDR6, DCL4 or DCL1 in a ddm1 background results in loss of 21-nucleotide easiRNAs and severe infertility, but 24-nucleotide hetsiRNAs are partially restored, supporting an antagonistic relationship between PTGS and TGS. Thus miRNA-directed easiRNA biogenesis is a latent mechanism that specifically targets transposon transcripts, but only when they are epigenetically reactivated during reprogramming of the germ line. This ancient recognition mechanism may have been retained both by transposons to evade long-term heterochromatic silencing and by their hosts for genome defence.
Figures

Comment in
-
RNA interference: microRNAs suppress transposons.Nat Rev Mol Cell Biol. 2014 May;15(5):298-9. doi: 10.1038/nrm3788. Epub 2014 Apr 3. Nat Rev Mol Cell Biol. 2014. PMID: 24694981 No abstract available.
-
Exploring new models of easiRNA biogenesis.Nat Genet. 2014 Jun;46(6):530-1. doi: 10.1038/ng.2993. Nat Genet. 2014. PMID: 24866189
References
-
- Allen E, Xie Z, Gustafson AM, Carrington JC. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
