

A novel tetravalent bispecific TandAb (CD30/CD16A) efficiently recruits NK cells for the lysis of CD30+ tumor cells
- PMID: 24670809
- PMCID: PMC4011917
- DOI: 10.4161/mabs.28591
A novel tetravalent bispecific TandAb (CD30/CD16A) efficiently recruits NK cells for the lysis of CD30+ tumor cells
Abstract
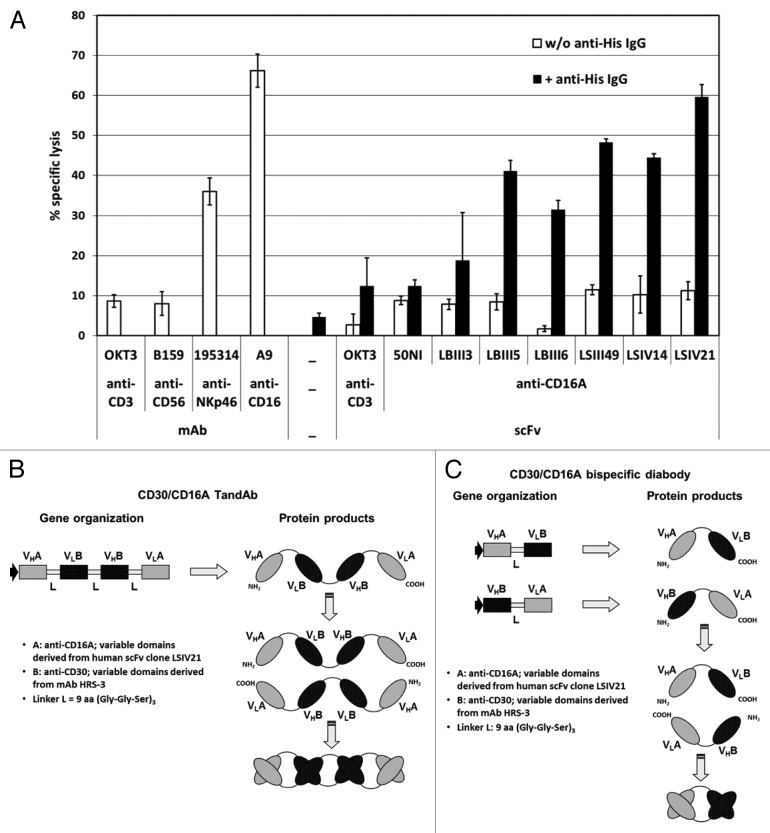
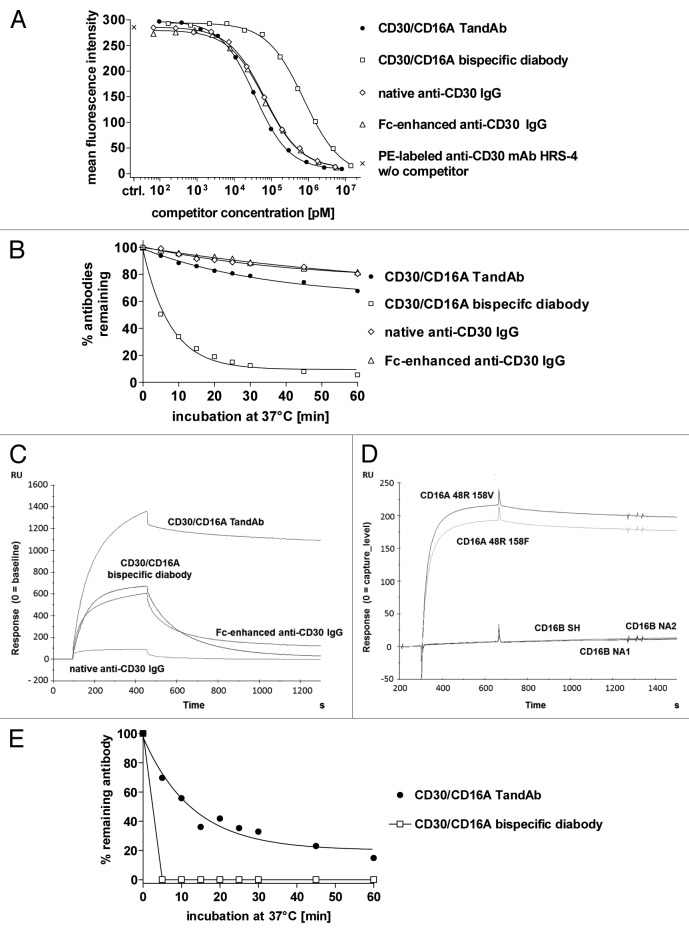
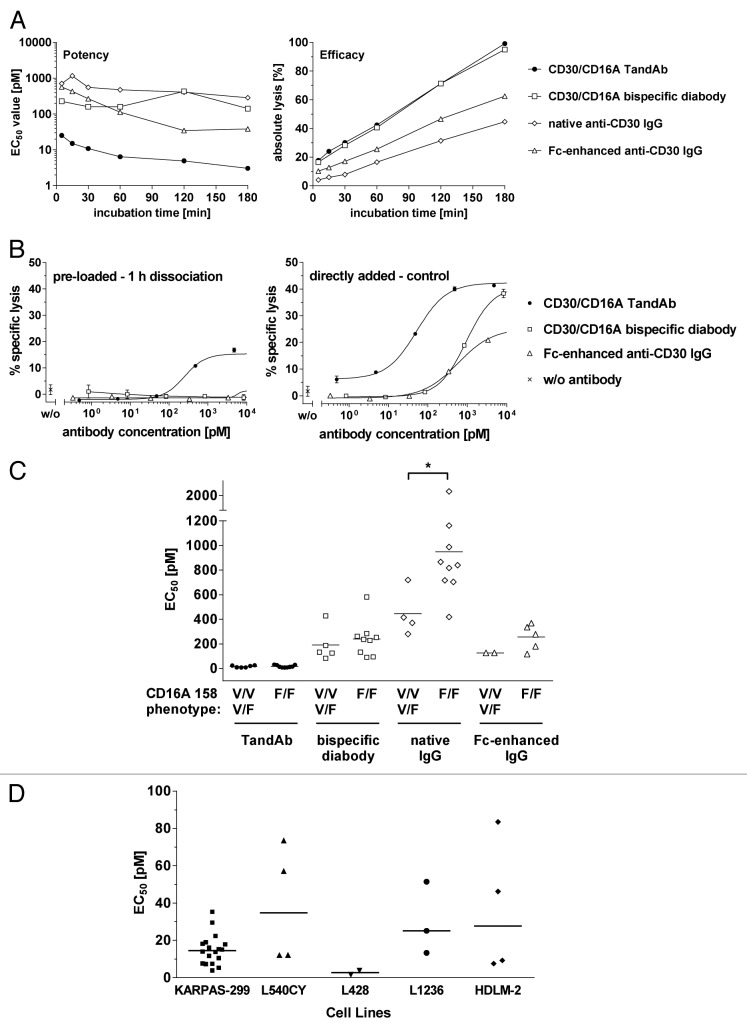
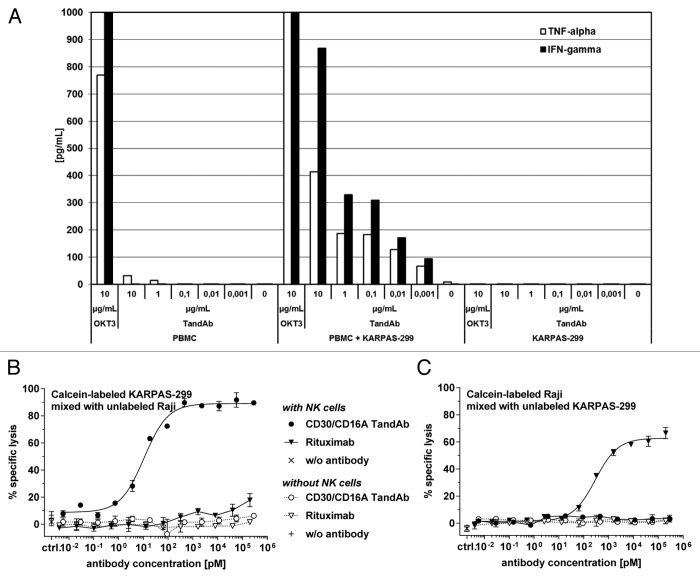
To improve recruitment and activation of natural killer (NK) cells to lyse tumor cells, we isolated a human anti-CD16A antibody with similar affinity for the CD16A 158F/V allotypes, but no binding to the CD16B isoform. Using CD16A-targeting Fv domains, we constructed a tetravalent bispecific CD30/CD16A tandem diabody (TandAb®) consisting solely of Fv domains. This TandAb has two binding sites for CD16A and two for CD30, the antigen identifying Hodgkin lymphoma cells. The binding and cytotoxicity of the TandAb were compared with antibodies with identical anti-CD30 domains: (1) a native IgG, (2) an IgG optimized for binding to Fc receptors, and (3) a bivalent bispecific CD30/CD16A diabody. Due to its CD16A-bivalency and reduced koff, the TandAb was retained longer on the surface of NK cells than the IgGs or the diabody. This contributed to the higher potency and efficacy of the TandAb relative to those of the other anti-CD30 antibodies. TandAb cytotoxicity was independent of the CD16A allotype, whereas the anti-CD30 IgGs were substantially less cytotoxic when NK cells with low affinity CD16A allotype were employed. TandAb activation of NK cells was strictly dependent on the presence of CD30(+) target cells. Therefore, the CD30/CD16A TandAb may represent a promising therapeutic for the treatment of Hodgkin's lymphoma; further, anti-CD16A TandAbs may function as potent immunotherapeutics that specifically recruit NK cells to destroy cancer cells.
Keywords: CD16A; FcγRIIIA; Hodgkin’s lymphoma; NK cells; bispecific antibodies.
Figures
References
-
- Robertson MJ, Ritz J. Biology and clinical relevance of human natural killer cells. Blood. 1990;76:2421–38. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases