

Gene and protein expression in response to different growth temperatures and oxygen availability in Burkholderia thailandensis
- PMID: 24671187
- PMCID: PMC3966863
- DOI: 10.1371/journal.pone.0093009
Gene and protein expression in response to different growth temperatures and oxygen availability in Burkholderia thailandensis
Abstract
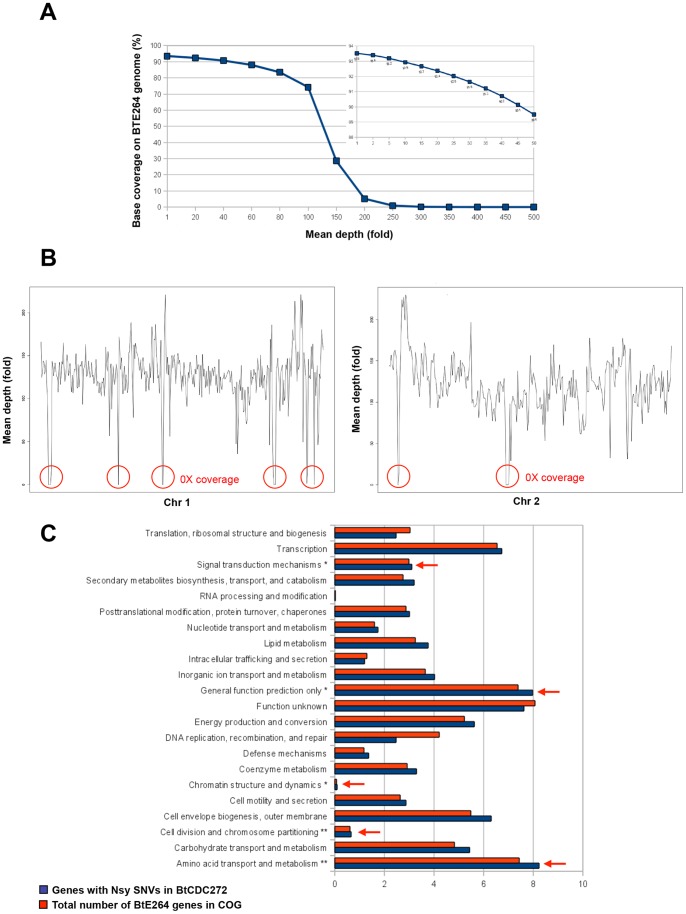
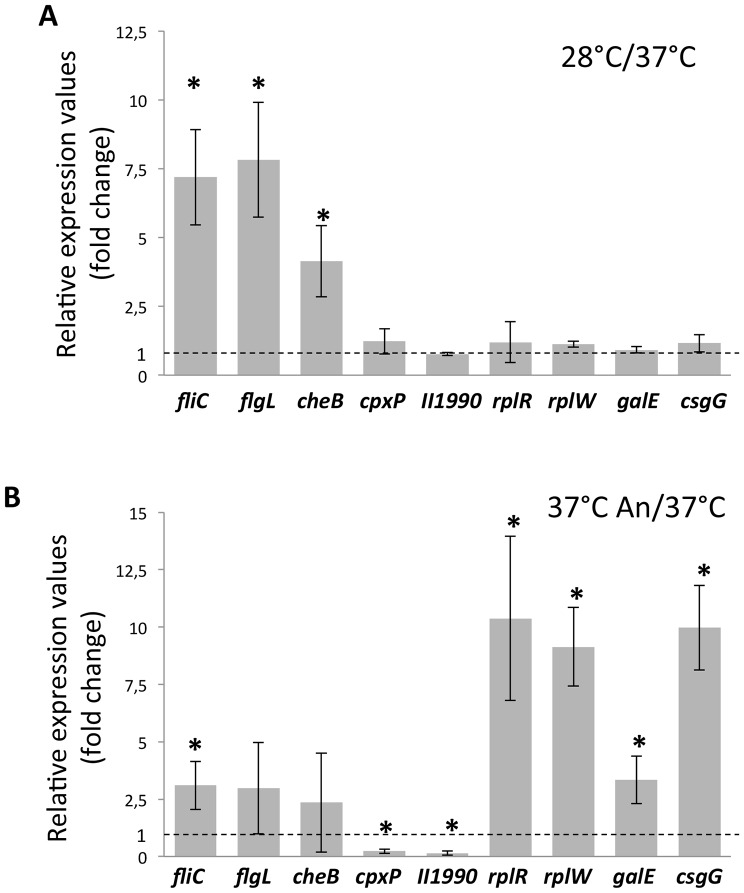
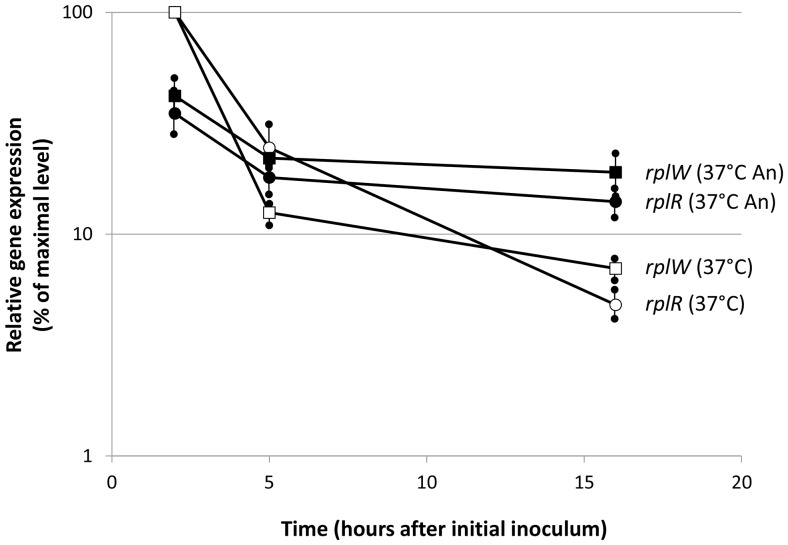
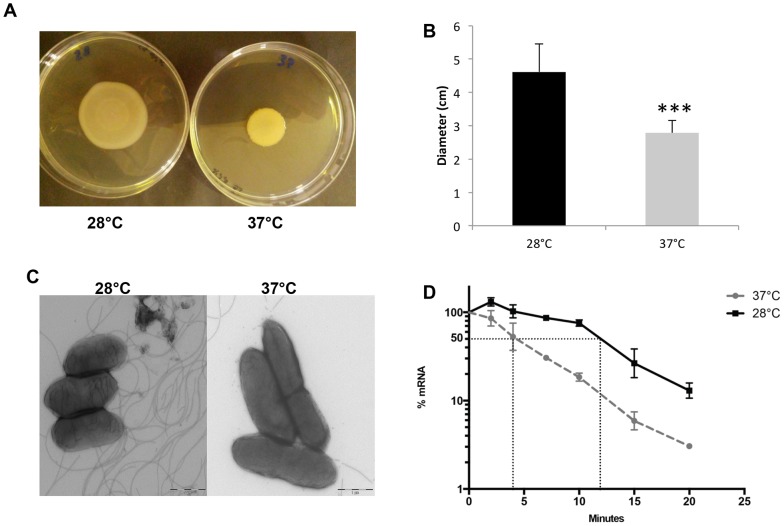
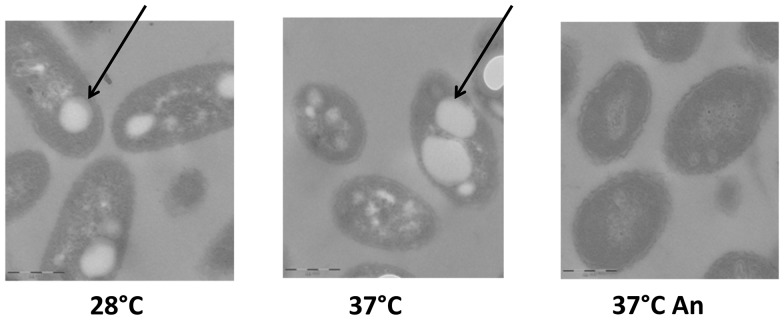
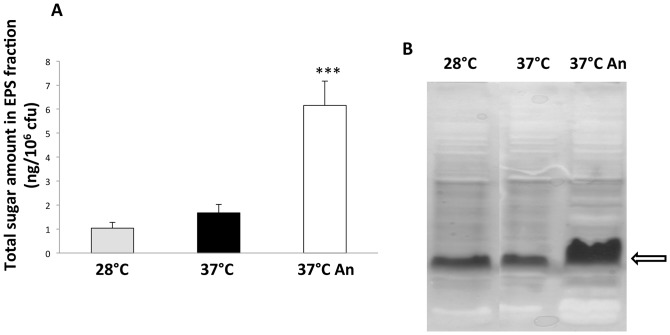
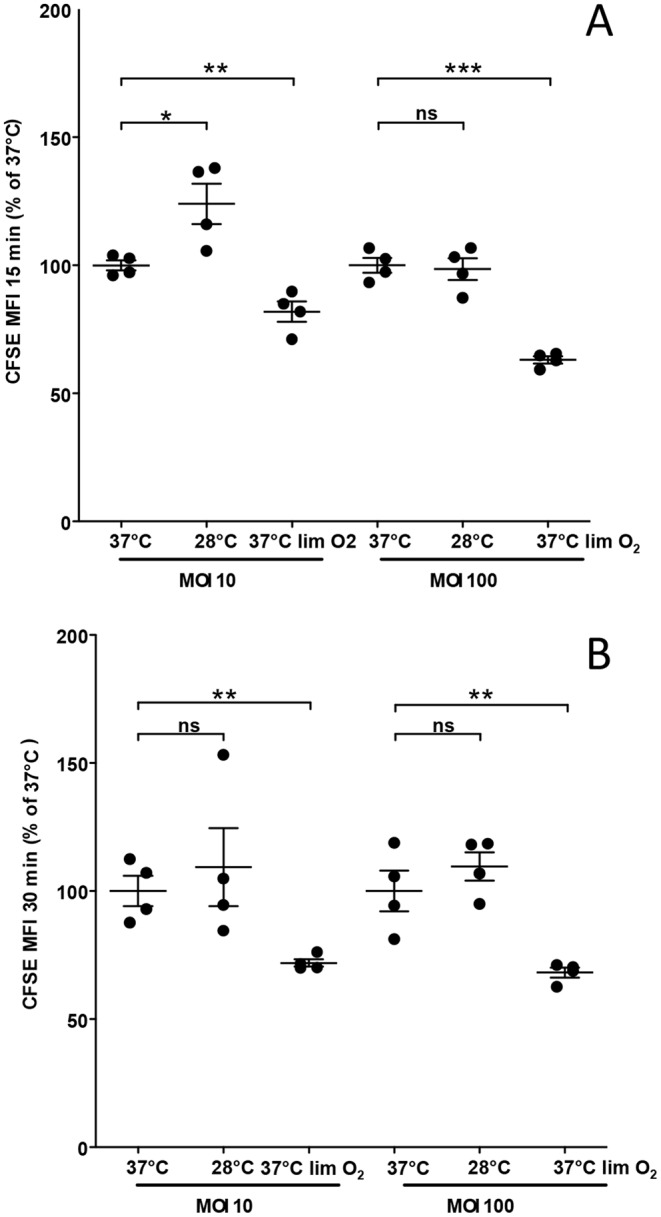
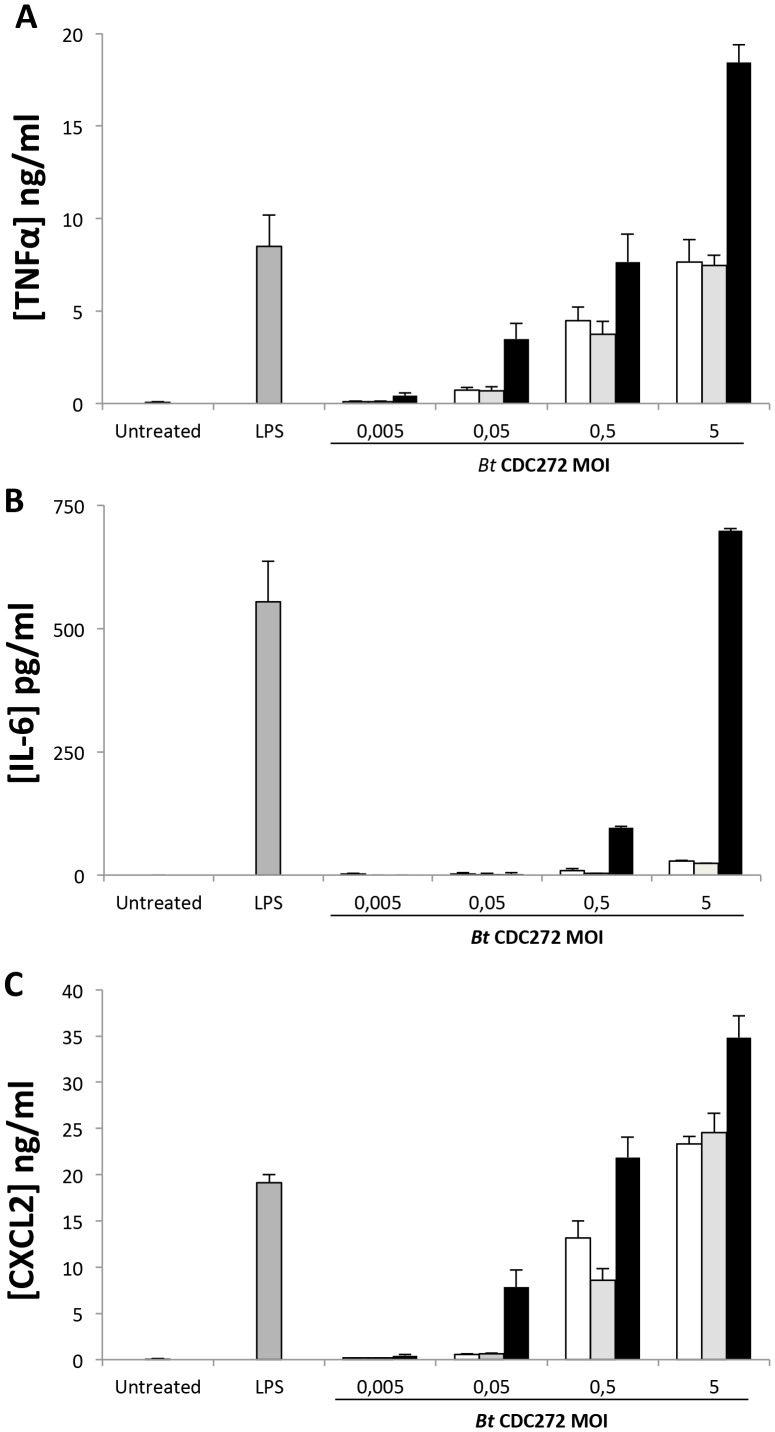
Burkholderia thailandensis, although normally avirulent for mammals, can infect macrophages in vitro and has occasionally been reported to cause pneumonia in humans. It is therefore used as a model organism for the human pathogen B. pseudomallei, to which it is closely related phylogenetically. We characterized the B. thailandensis clinical isolate CDC2721121 (BtCDC272) at the genome level and studied its response to environmental cues associated with human host colonization, namely, temperature and oxygen limitation. Effects of the different growth conditions on BtCDC272 were studied through whole genome transcription studies and analysis of proteins associated with the bacterial cell surface. We found that growth at 37°C, compared to 28°C, negatively affected cell motility and flagella production through a mechanism involving regulation of the flagellin-encoding fliC gene at the mRNA stability level. Growth in oxygen-limiting conditions, in contrast, stimulated various processes linked to virulence, such as lipopolysaccharide production and expression of genes encoding protein secretion systems. Consistent with these observations, BtCDC272 grown in oxygen limitation was more resistant to phagocytosis and strongly induced the production of inflammatory cytokines from murine macrophages. Our results suggest that, while temperature sensing is important for regulation of B. thailandensis cell motility, oxygen limitation has a deeper impact on its physiology and constitutes a crucial environmental signal for the production of virulence factors.
Conflict of interest statement
Figures
References
-
- Coenye T, Vandamme P (2003) Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ Microbiol 5: 719–729. - PubMed
-
- Mahenthiralingam E, Urban TA, Goldberg JB (2005) The multifarious, multireplicon Burkholderia cepacia complex. Nat Rev Microbiol 3: 144–156. - PubMed
-
- White NJ (2003) Melioidosis. Lancet 361: 1715–1722. - PubMed
-
- Brett PJ, DeShazer D, Woods DE (1998) Burkholderia thailandensis sp. nov., a Burkholderia pseudomallei-like species. Int J Syst Bacteriol 48 Pt 1: 317–320. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
