

Structural characterization of heparin-induced glyceraldehyde-3-phosphate dehydrogenase protofibrils preventing α-synuclein oligomeric species toxicity
- PMID: 24671416
- PMCID: PMC4022857
- DOI: 10.1074/jbc.M113.544288
Structural characterization of heparin-induced glyceraldehyde-3-phosphate dehydrogenase protofibrils preventing α-synuclein oligomeric species toxicity
Abstract
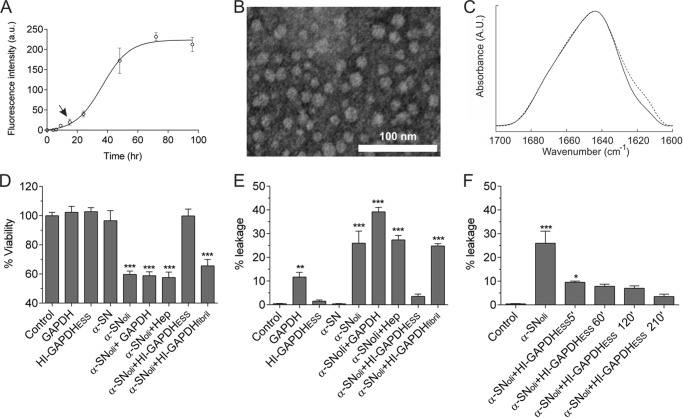
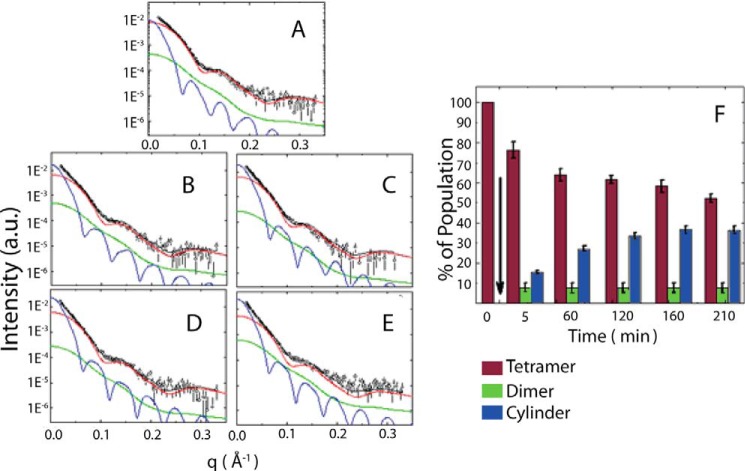
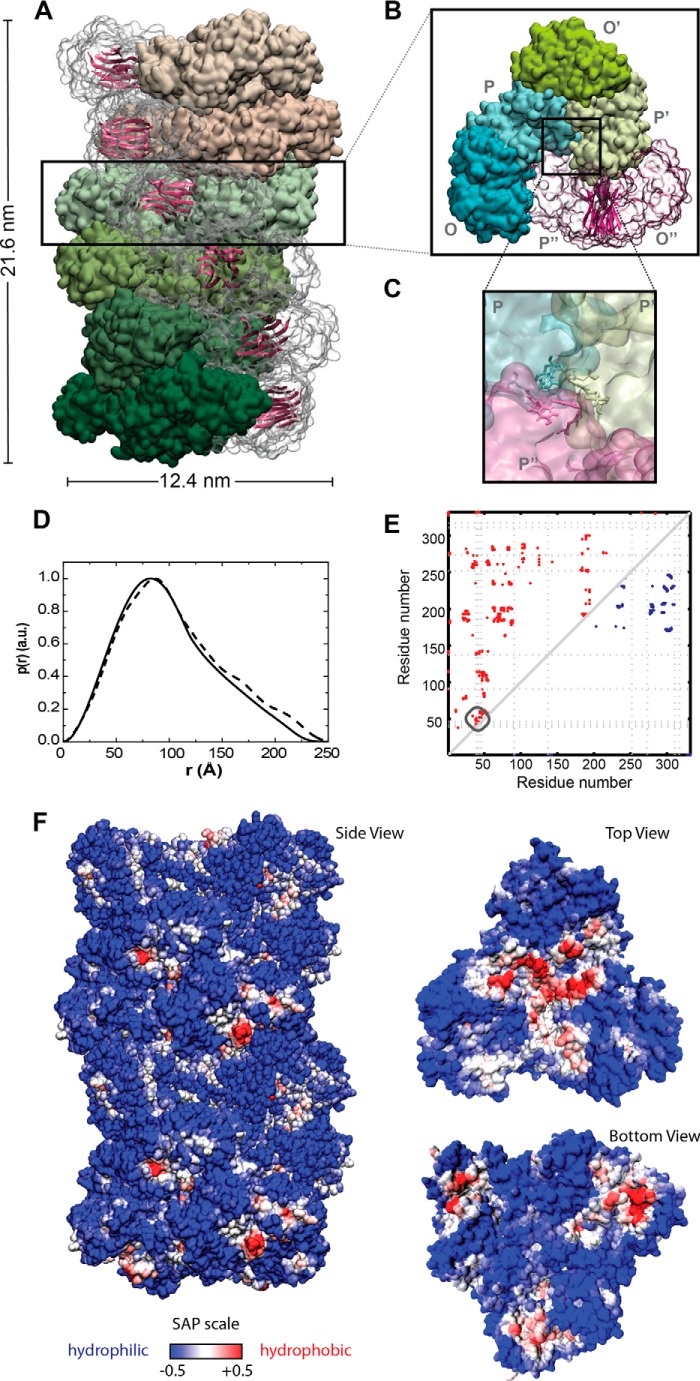
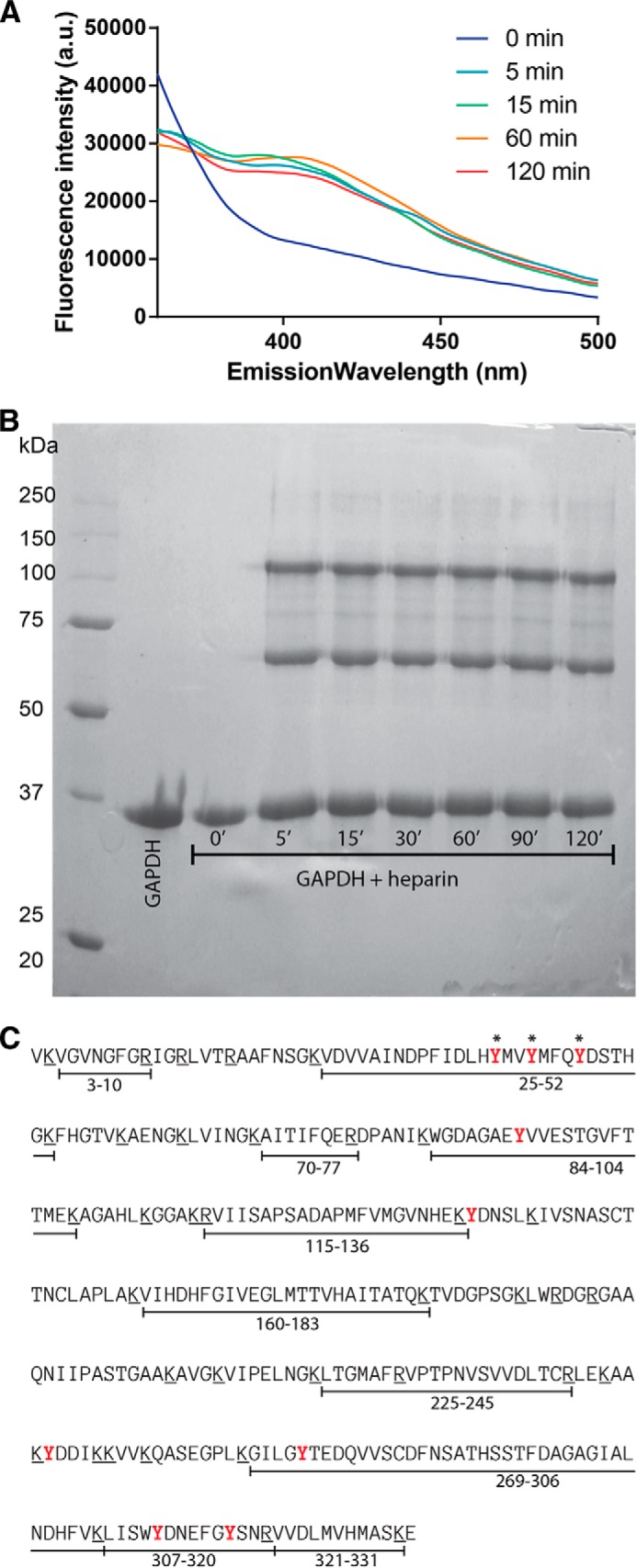
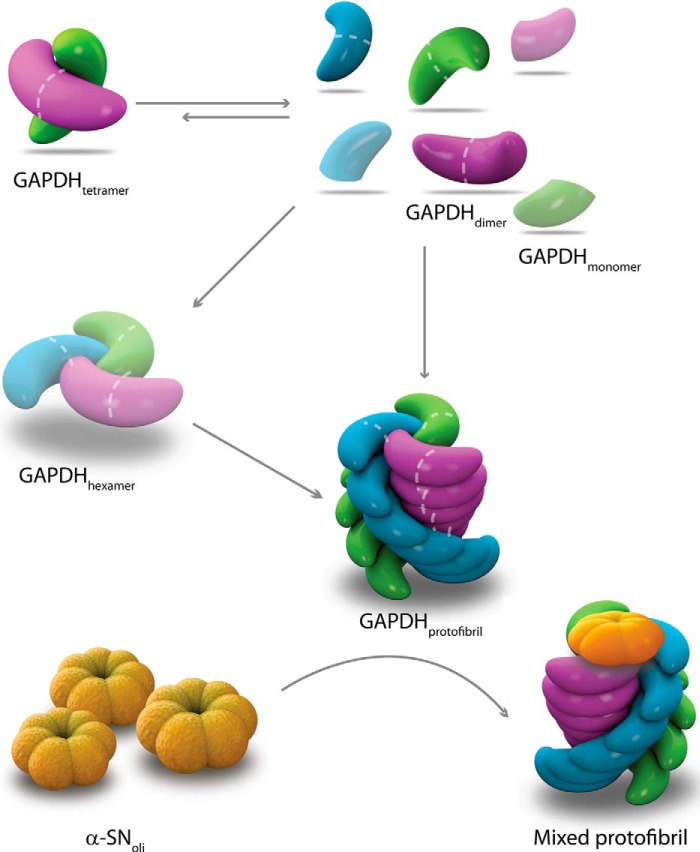
Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) is a multifunctional enzyme that has been associated with neurodegenerative diseases. GAPDH colocalizes with α-synuclein in amyloid aggregates in post-mortem tissue of patients with sporadic Parkinson disease and promotes the formation of Lewy body-like inclusions in cell culture. In a previous work, we showed that glycosaminoglycan-induced GAPDH prefibrillar species accelerate the conversion of α-synuclein to fibrils. However, it remains to be determined whether the interplay among glycosaminoglycans, GAPDH, and α-synuclein has a role in pathological states. Here, we demonstrate that the toxic effect exerted by α-synuclein oligomers in dopaminergic cell culture is abolished in the presence of GAPDH prefibrillar species. Structural analysis of prefibrillar GAPDH performed by small angle x-ray scattering showed a particle compatible with a protofibril. This protofibril is shaped as a cylinder 22 nm long and a cross-section diameter of 12 nm. Using biocomputational techniques, we obtained the first all-atom model of the GAPDH protofibril, which was validated by cross-linking coupled to mass spectrometry experiments. Because GAPDH can be secreted outside the cell where glycosaminoglycans are present, it seems plausible that GAPDH protofibrils could be assembled in the extracellular space kidnapping α-synuclein toxic oligomers. Thus, the role of GAPDH protofibrils in neuronal proteostasis must be considered. The data reported here could open alternative ways in the development of therapeutic strategies against synucleinopathies like Parkinson disease.
Keywords: Cell Permeabilization; GAPDH; Molecular Modeling; Parkinson Disease; Protofibril Structure; X-ray Scattering; α-Synuclein.
© 2014 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Lee H. J., Suk J. E., Bae E. J., Lee S. J. (2008) Clearance and deposition of extracellular α-synuclein aggregates in microglia. Biochem. Biophys. Res. Commun. 372, 423–428 - PubMed
-
- Lee S. J. (2008) Origins and effects of extracellular α-synuclein: implications in Parkinson's disease. J. Mol. Neurosci. 34, 17–22 - PubMed
-
- Chai Y. J., Kim D., Park J., Zhao H., Lee S. J., Chang S. (2013) The secreted oligomeric form of α-synuclein affects multiple steps of membrane trafficking. FEBS Lett. 587, 452–459 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
