

Biophysics of directional hearing in the American alligator (Alligator mississippiensis)
- PMID: 24671963
- PMCID: PMC3966920
- DOI: 10.1242/jeb.092866
Biophysics of directional hearing in the American alligator (Alligator mississippiensis)
Abstract
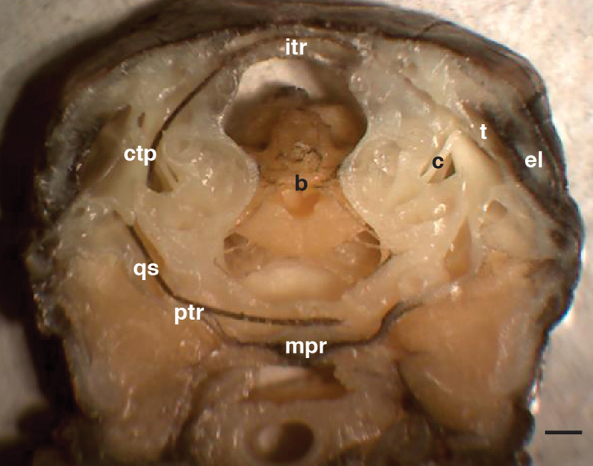
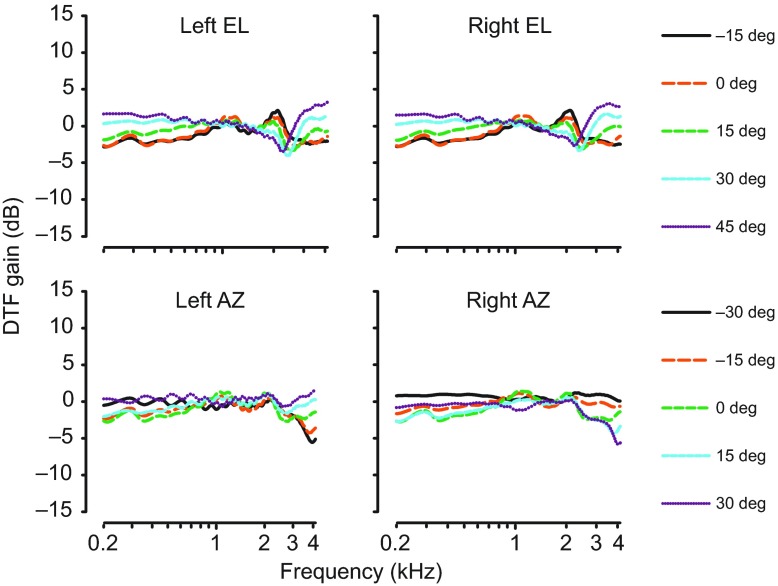
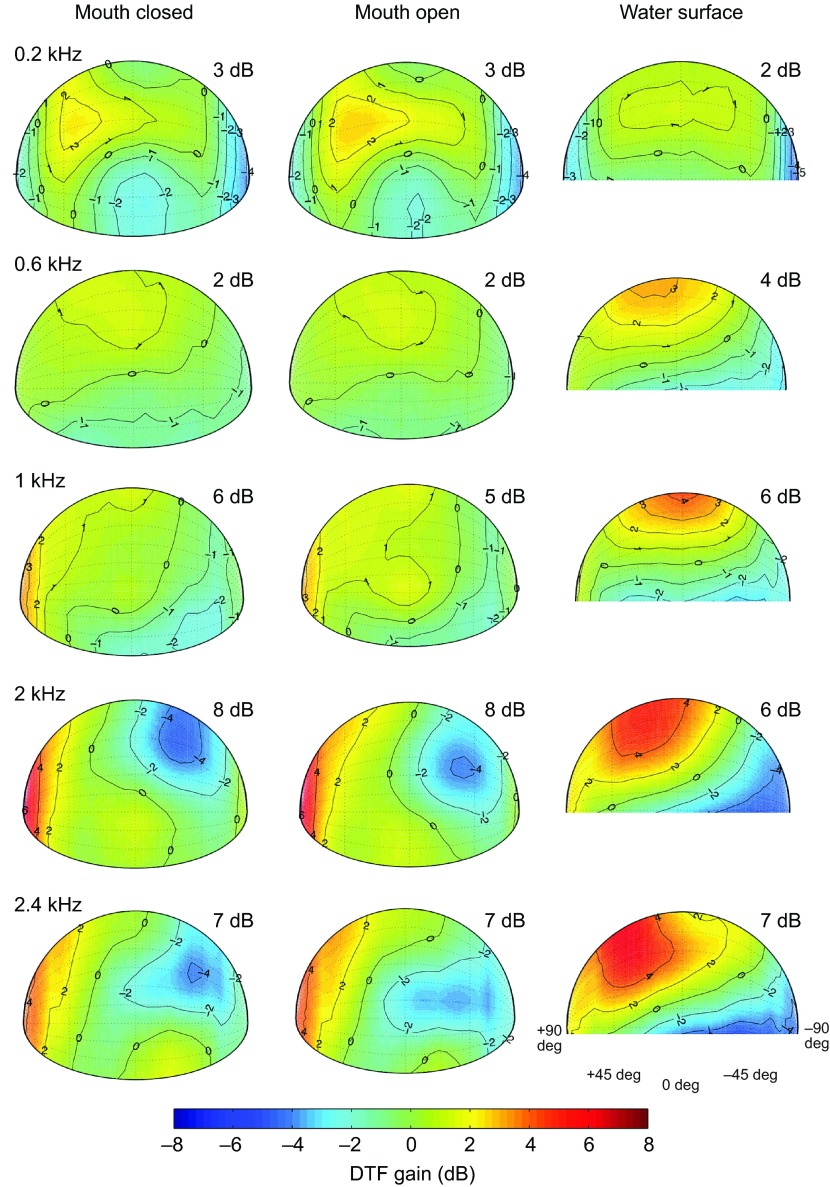
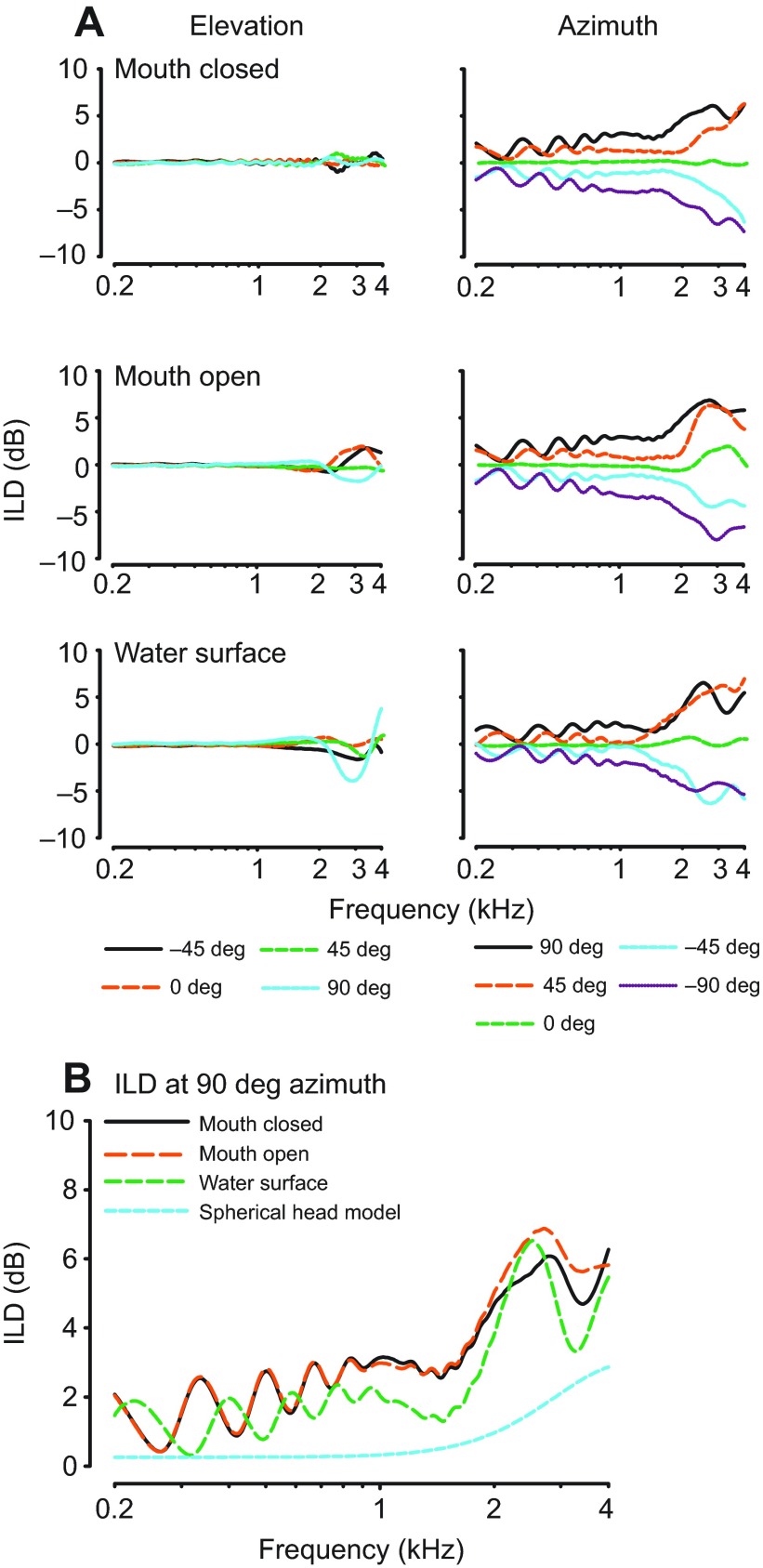
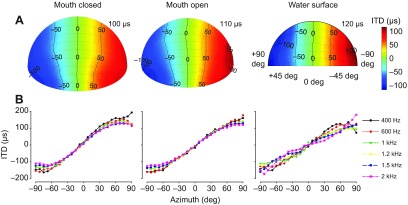
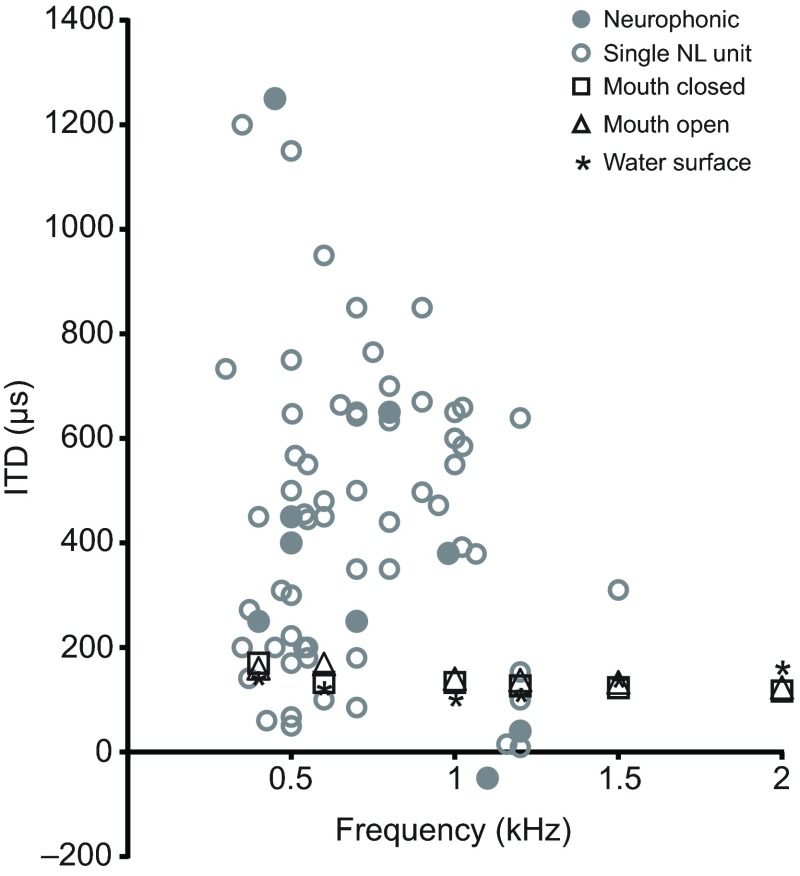
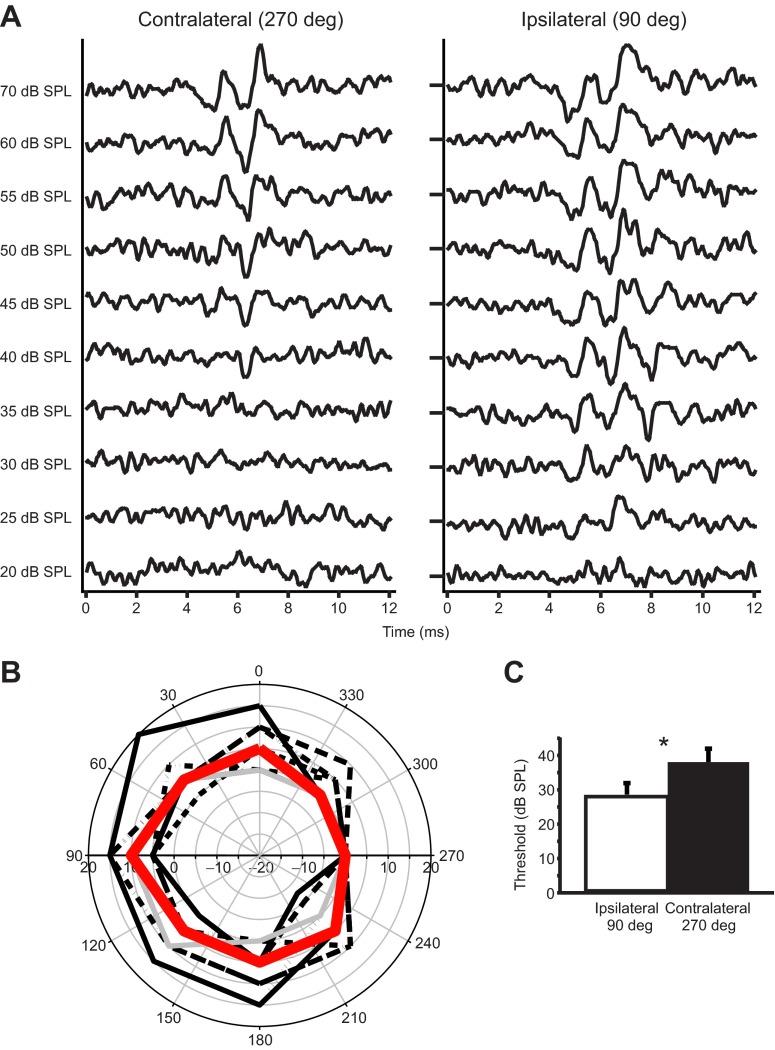
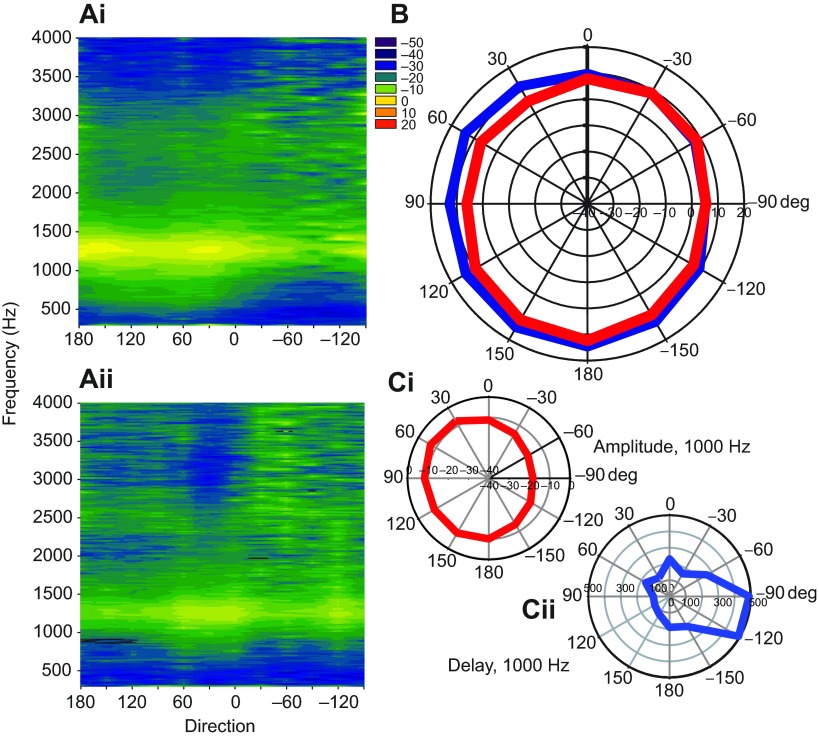
Physiological and anatomical studies have suggested that alligators have unique adaptations for spatial hearing. Sound localization cues are primarily generated by the filtering of sound waves by the head. Different vertebrate lineages have evolved external and/or internal anatomical adaptations to enhance these cues, such as pinnae and interaural canals. It has been hypothesized that in alligators, directionality may be enhanced via the acoustic coupling of middle ear cavities, resulting in a pressure difference receiver (PDR) mechanism. The experiments reported here support a role for a PDR mechanism in alligator sound localization by demonstrating that (1) acoustic space cues generated by the external morphology of the animal are not sufficient to generate location cues that match physiological sensitivity, (2) continuous pathways between the middle ears are present to provide an anatomical basis for coupling, (3) the auditory brainstem response shows some directionality, and (4) eardrum movement is directionally sensitive. Together, these data support the role of a PDR mechanism in crocodilians and further suggest this mechanism is a shared archosaur trait, most likely found also in the extinct dinosaurs.
Keywords: ABR; Archosaur; Auditory; Bird; Dinosaur; HRTF; Middle ear; Pressure-receiver; Vibrometry.
Figures

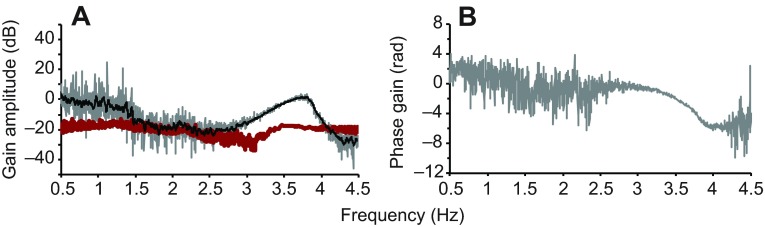
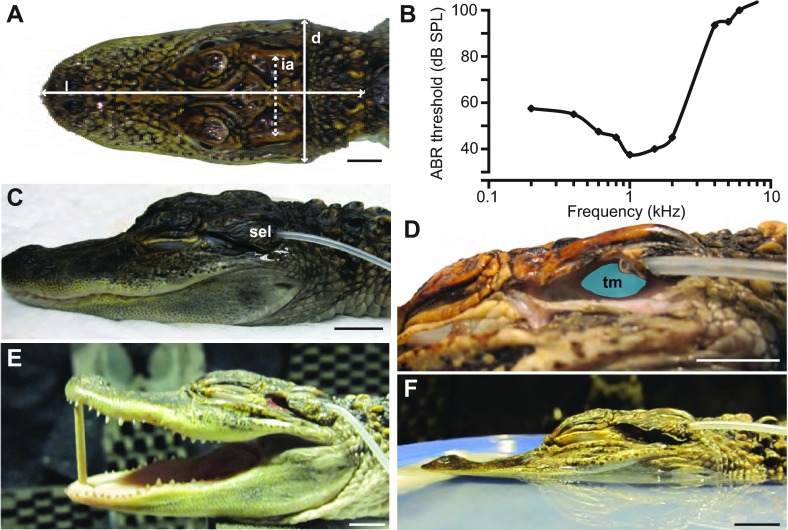
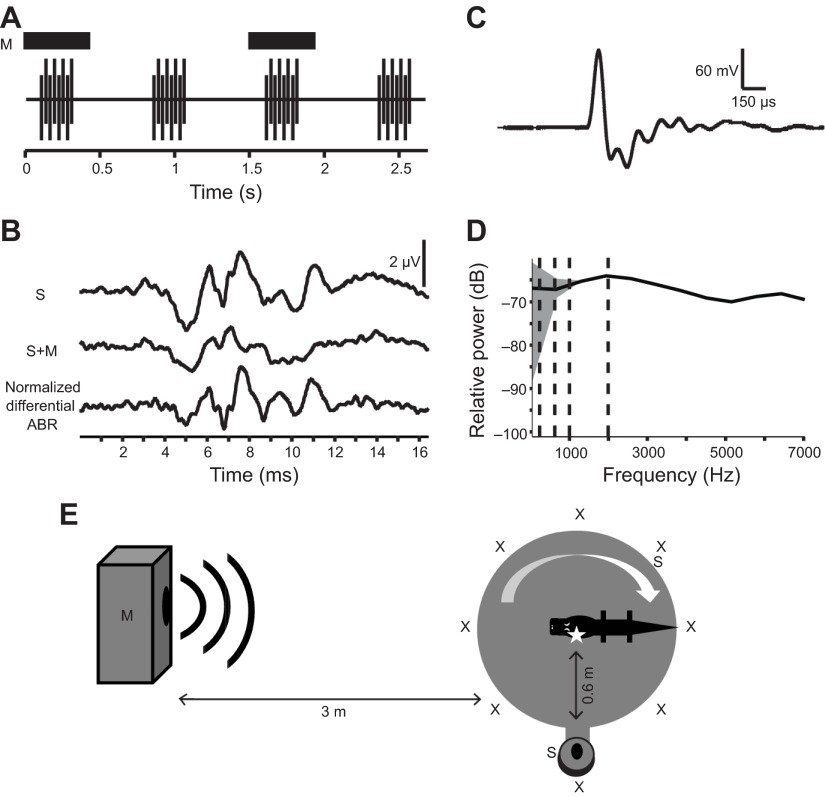
References
-
- Autrum H. (1940). Über lautäusserungen und schallwahrnehmung bei arthropoden II. Z. Vgl Physiol. 28, 326-352
-
- Aytekin M., Grassi E., Sahota M., Moss C. F. (2004). The bat head-related transfer function reveals binaural cues for sound localization in azimuth and elevation. J. Acoust. Soc. Am. 116, 3594-3605 - PubMed
-
- Batra R., Kuwada S., Fitzpatrick D. C. (1997). Sensitivity to interaural temporal disparities of low- and high-frequency neurons in the superior olivary complex. I. Heterogeneity of responses. J. Neurophysiol. 78, 1222-1236 - PubMed
-
- Bierman H. S., Carr C. E., Brandt C., Young B. A., Christensen-Dalsgaard J. (2011). Evidence for low-frequency sound localization in the American alligator (Alligator mississippiensis). In 2011 Neuroscience Meeting Planner. Washington, DC: Society for Neuroscience. Online;
-
- Boettcher F. A., Mills J. H., Norton B. L. (1993). Age-related changes in auditory evoked potentials of gerbils. I. Response amplitudes. Hear. Res. 71, 137-145 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
