

Human metapneumovirus virus-like particles induce protective B and T cell responses in a mouse model
- PMID: 24672031
- PMCID: PMC4093842
- DOI: 10.1128/JVI.00332-14
Human metapneumovirus virus-like particles induce protective B and T cell responses in a mouse model
Abstract
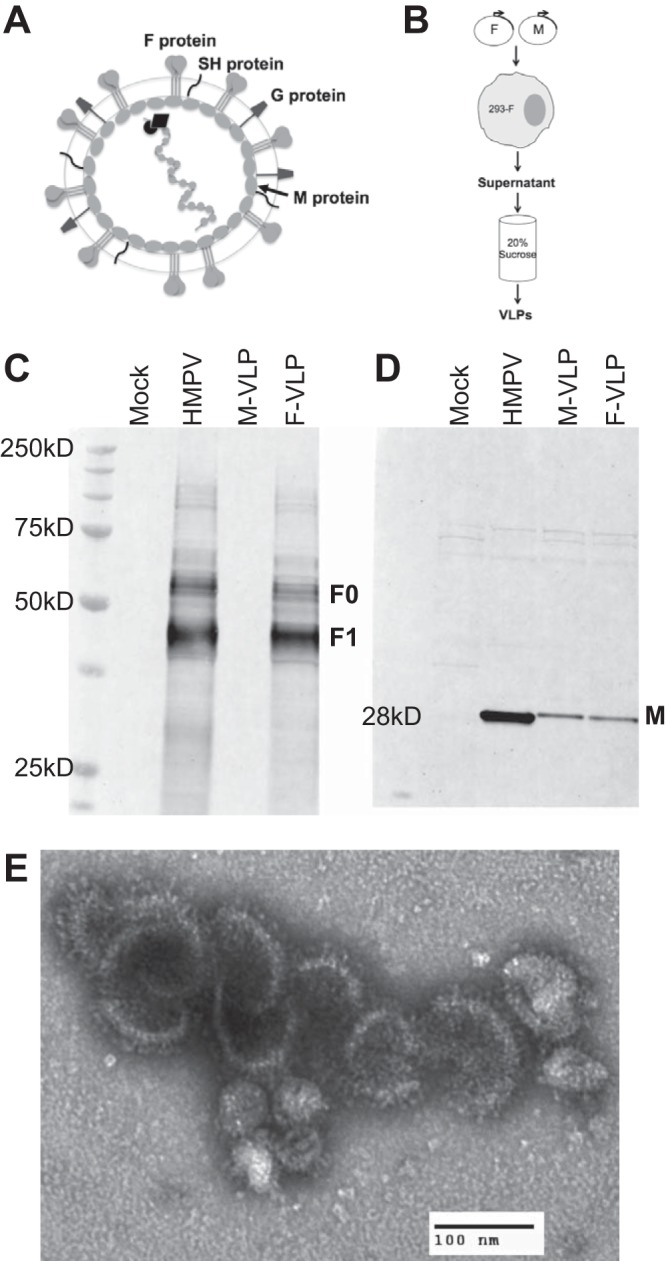
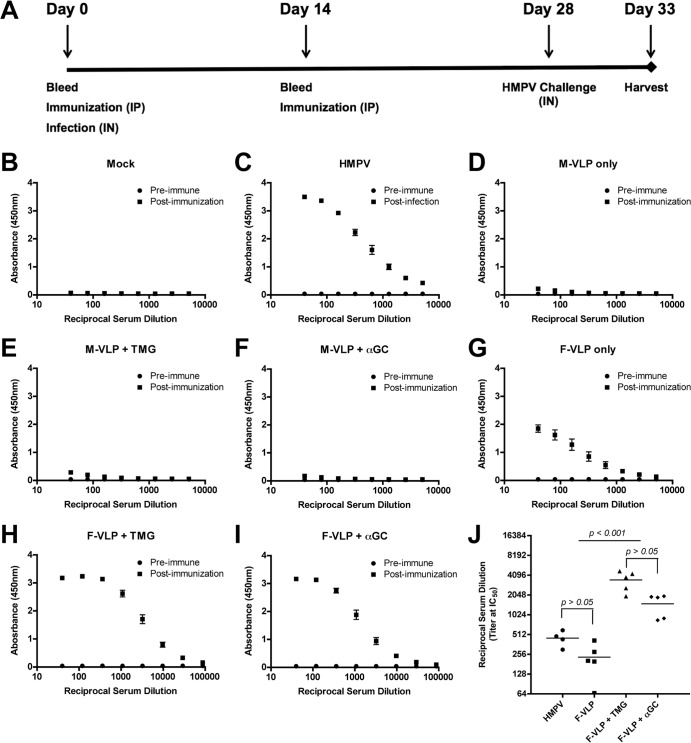
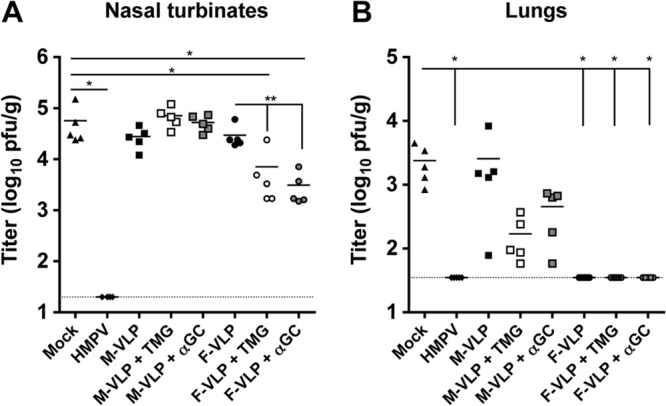
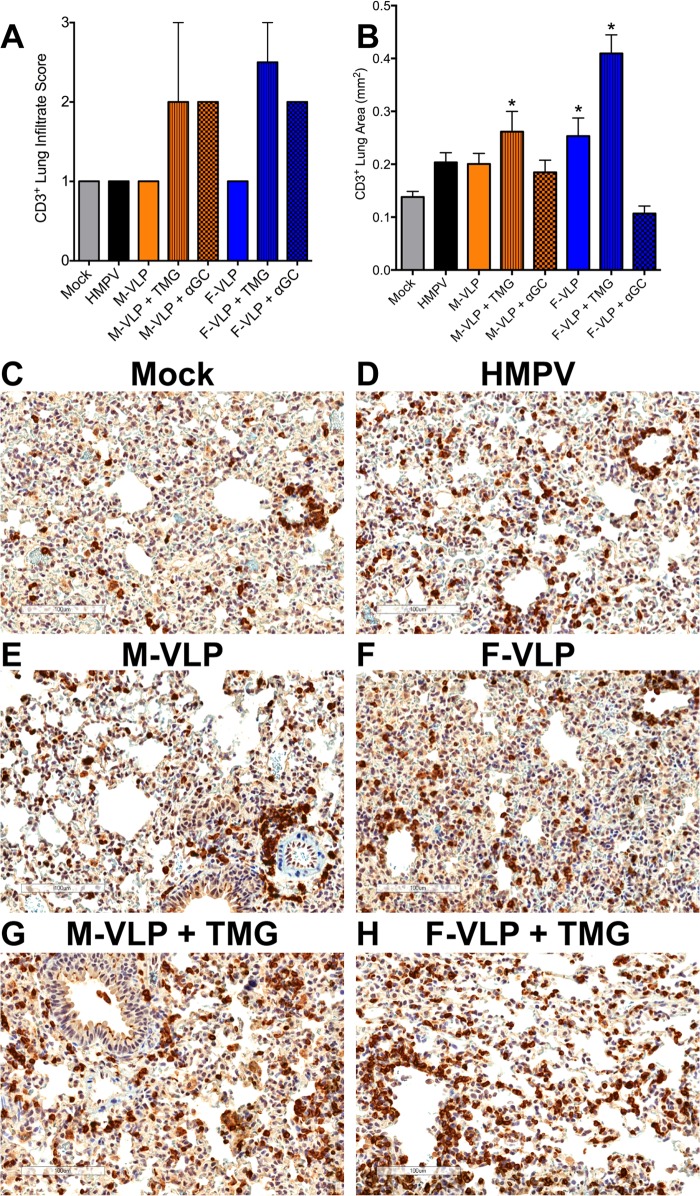
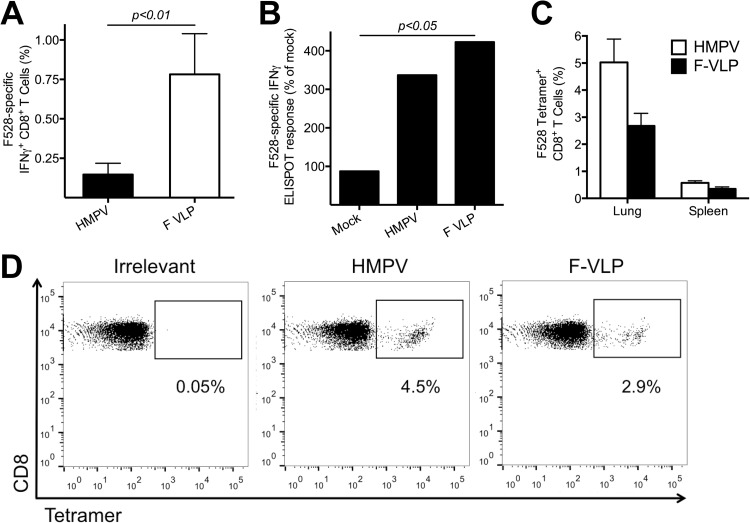
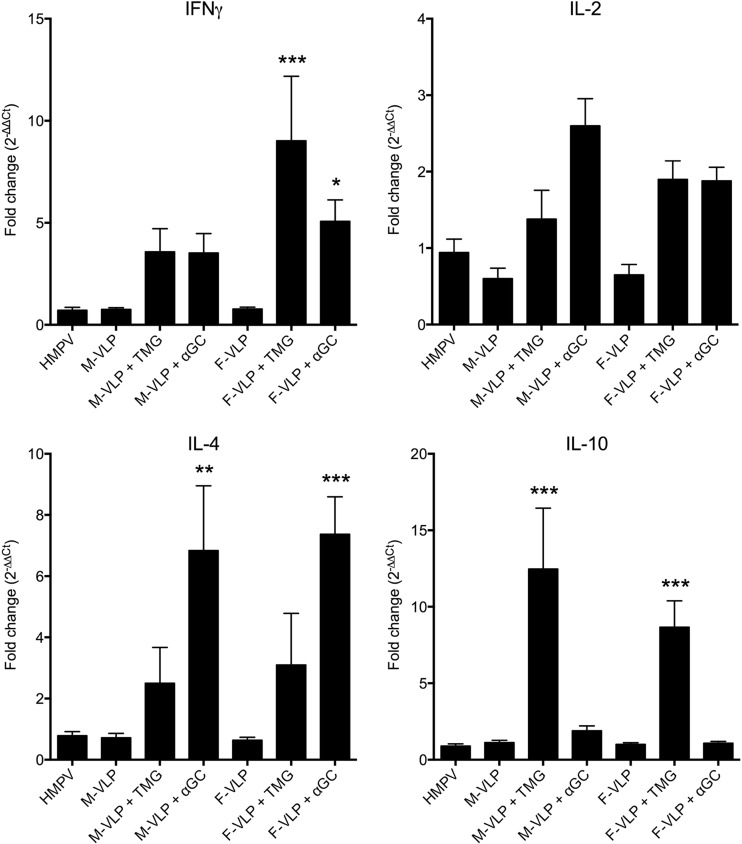
Human metapneumovirus (HMPV) is a leading cause of respiratory disease in infants, children, and the elderly worldwide, yet no licensed vaccines exist. Live-attenuated vaccines present safety challenges, and protein subunit vaccines induce primarily antibody responses. Virus-like particles (VLPs) are an attractive alternative vaccine approach because of reduced safety concerns compared with live vaccines. We generated HMPV VLPs by expressing viral proteins in suspension-adapted human embryonic kidney epithelial (293-F) cells and found that the viral matrix (M) and fusion (F) proteins were sufficient to form VLPs. We previously reported that the VLPs resemble virus morphology and incorporate fusion-competent F protein (R. G. Cox, S. B. Livesay, M. Johnson, M. D. Ohi, and J. V. Williams, J. Virol. 86:12148-12160, 2012), which we hypothesized would elicit F-specific antibody and T cell responses. In this study, we tested whether VLP immunization could induce protective immunity to HMPV by using a mouse model. C57BL/6 mice were injected twice intraperitoneally with VLPs alone or with adjuvant and subsequently challenged with HMPV. Mice were euthanized 5 days postinfection, and virus titers, levels of neutralizing antibodies, and numbers of CD3(+) T cells were quantified. Mice immunized with VLPs mounted an F-specific antibody response and generated CD8(+) T cells recognizing an F protein-derived epitope. VLP immunization induced a neutralizing-antibody response that was enhanced by the addition of either TiterMax Gold or α-galactosylceramide adjuvant, though adjuvant reduced cellular immune responses. Two doses of VLPs conferred complete protection from HMPV replication in the lungs of mice and were not associated with a Th2-skewed cytokine response. These results suggest that nonreplicating VLPs are a promising vaccine candidate for HMPV.
Importance: Human metapneumovirus (HMPV) is a leading cause of acute respiratory infection in infants, children, and the elderly worldwide, yet no licensed vaccines exist. Live-attenuated vaccines present safety challenges, and protein subunit vaccines induce primarily antibody responses. Virus-like particles (VLPs) are an attractive alternative vaccine approach. We generated HMPV VLPs by expressing the viral matrix (M) and fusion (F) proteins in mammalian cells. We found that mice immunized with VLPs mounted an F-specific antibody response and generated CD8(+) T cells recognizing an F protein-derived epitope. VLP immunization induced a neutralizing-antibody response that was enhanced by the addition of either TiterMax Gold or α-galactosylceramide adjuvant. Two doses of VLPs conferred complete protection against HMPV replication in the lungs of mice and were not associated with a Th2-skewed cytokine response. These results suggest that nonreplicating VLPs are a promising vaccine candidate for HMPV.
Figures
References
-
- Williams JV, Wang CK, Yang CF, Tollefson SJ, House FS, Heck JM, Chu M, Brown JB, Lintao LD, Quinto JD, Chu D, Spaete RR, Edwards KM, Wright PF, Crowe JE., Jr 2006. The role of human metapneumovirus in upper respiratory tract infections in children: a 20-year experience. J. Infect. Dis. 193:387–395. 10.1086/499274 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- P60 DK020593/DK/NIDDK NIH HHS/United States
- R01 AI085062/AI/NIAID NIH HHS/United States
- GM-007347/GM/NIGMS NIH HHS/United States
- T32 AI-007611/AI/NIAID NIH HHS/United States
- T32 HL069765/HL/NHLBI NIH HHS/United States
- P30 CA068485/CA/NCI NIH HHS/United States
- P60 DK-020593/DK/NIDDK NIH HHS/United States
- AI-085062/AI/NIAID NIH HHS/United States
- P30 CA-68485/CA/NCI NIH HHS/United States
- P30 DK058404/DK/NIDDK NIH HHS/United States
- T32 HL-069765/HL/NHLBI NIH HHS/United States
- T32 GM007347/GM/NIGMS NIH HHS/United States
- P30 DK020593/DK/NIDDK NIH HHS/United States
- T32 AI007611/AI/NIAID NIH HHS/United States
- DK-058404/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
