

The Rad6/18 ubiquitin complex interacts with the Epstein-Barr virus deubiquitinating enzyme, BPLF1, and contributes to virus infectivity
- PMID: 24672041
- PMCID: PMC4093873
- DOI: 10.1128/JVI.00536-14
The Rad6/18 ubiquitin complex interacts with the Epstein-Barr virus deubiquitinating enzyme, BPLF1, and contributes to virus infectivity
Abstract
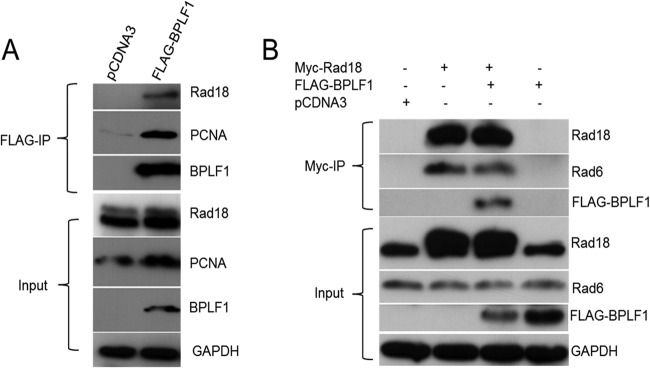
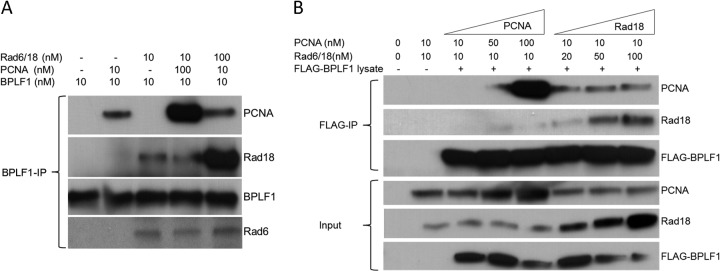
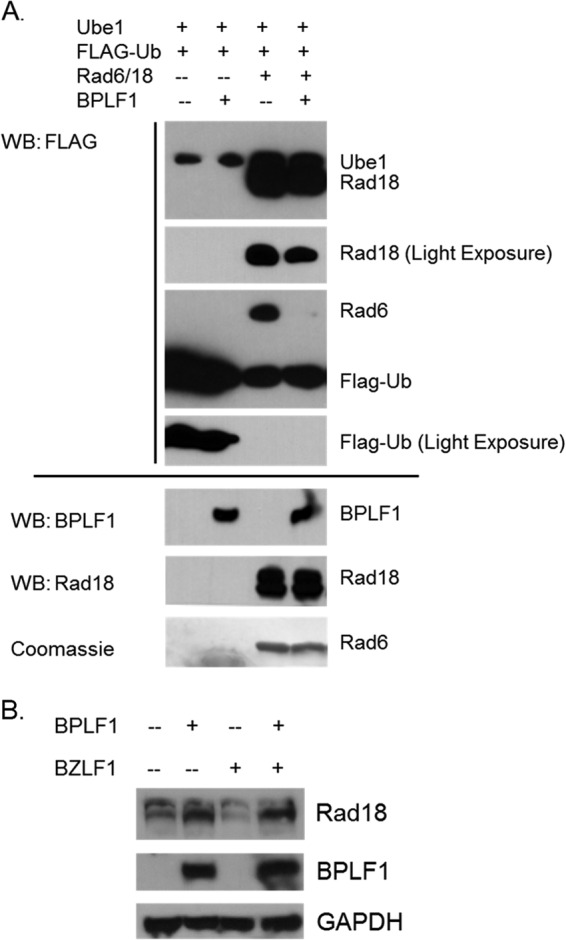
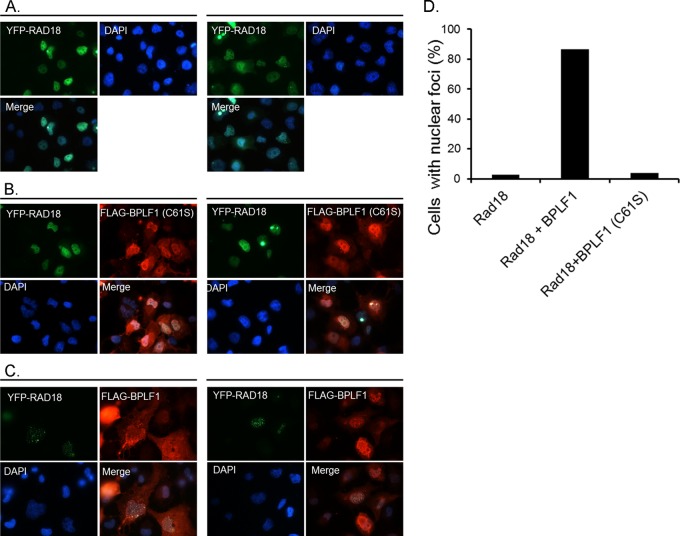
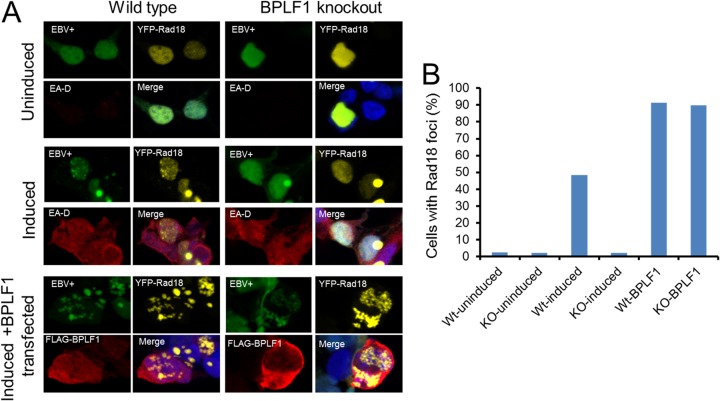
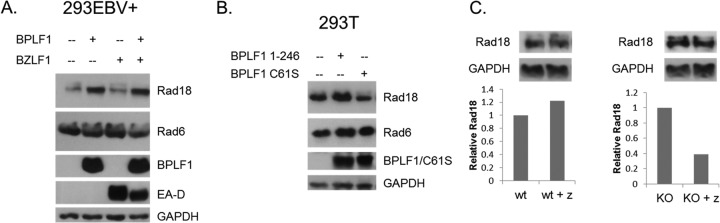
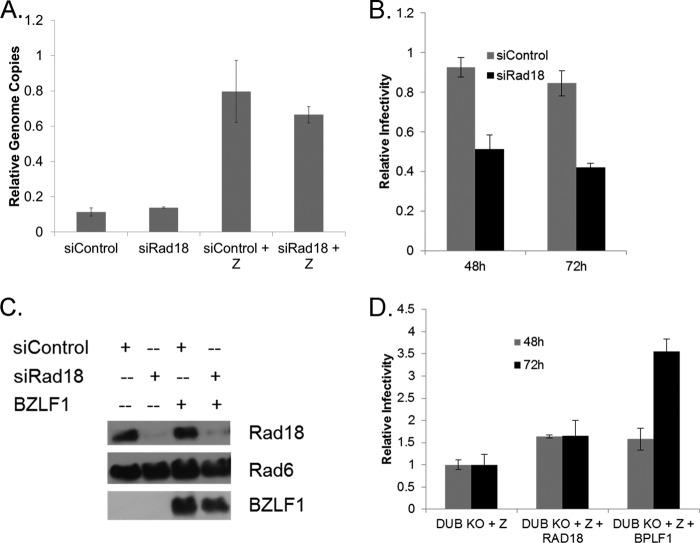
Epstein-Barr virus (EBV) encodes BPLF1, a lytic cycle protein with deubiquitinating activity that is contained in its N-terminal domain and conserved across the Herpesviridae. EBV replication is associated with cellular DNA replication and repair factors, and initiation of EBV lytic replication induces a DNA damage response, which can be regulated at least in part by BPLF1. The cellular DNA repair pathway, translesion synthesis (TLS), is disrupted by BPLF1, which deubiquitinates the DNA processivity factor, PCNA, and inhibits the recruitment of the TLS polymerase, polymerase eta (Pol eta), after damage to DNA by UV irradiation. Here we showed that the E3 ubiquitin ligase, which activates TLS repair by monoubiquitination of PCNA, is also affected by BPLF1 deubiquitinating activity. First, BPLF1 interacts directly with Rad18, and overexpression of BPLF1 results in increased levels of the Rad18 protein, suggesting that it stabilizes Rad18. Next, expression of functionally active BPLF1 caused relocalization of Rad18 into nuclear foci, which is consistent with sites of cellular DNA replication that occur during S phase. Also, levels of Rad18 remain constant during lytic reactivation of wild-type virus, but reactivation of BPLF1 knockout virus resulted in decreased levels of Rad18. Finally, the contribution of Rad18 levels to infectious virus production was examined with small interfering RNA (siRNA) targeting Rad18. Results demonstrated that reducing levels of Rad18 decreased production of infectious virus, and infectious titers of BPLF1 knockout virus were partially restored by overexpression of Rad18. Thus, BPLF1 interacts with and maintains Rad18 at high levels during lytic replication, which assists in production of infectious virus.
Importance: Characterization of EBV BPLF1's deubiquitinating activity and identification of its targets and subsequent functional effects remain little studied. All members of the Herpesviridae contain BPLF1 homologs with conserved enzymatic activity, and findings discovered with EBV BPLF1 are likely applicable to other members of the family. Discovery of new targets of BPLF1 will point to cellular pathways and viral processes regulated by the enzymatic activity of the EBV-encoded deubiquitinating enzyme. Here we determined the importance of the cellular ubiquitin ligase Rad18 in these processes and how it is affected by BPLF1. Our findings demonstrate that EBV can co-opt Rad18 as a novel accessory factor in the production of infectious virus.
Figures
References
-
- Anagnostopoulos I, Hummel M, Kreschel C, Stein H. 1995. Morphology, immunophenotype, and distribution of latently and/or productively Epstein-Barr virus-infected cells in acute infectious mononucleosis: implications for the interindividual infection route of Epstein-Barr virus. Blood 85:744–750 - PubMed
-
- Niederman JC, Miller G, Pearson HA, Pagano JS, Dowaliby JM. 1976. Infectious mononucleosis. Epstein-Barr-virus shedding in saliva and the oropharynx. N. Engl. J. Med. 294:1355–1359 - PubMed
-
- Pagano J. 2009. EBV diseases, p 794 In Damania B, Pipas J. (ed), DNA tumor viruses. Springer, New York, NY
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
