

LMTK1 regulates dendritic formation by regulating movement of Rab11A-positive endosomes
- PMID: 24672056
- PMCID: PMC4038502
- DOI: 10.1091/mbc.E14-01-0675
LMTK1 regulates dendritic formation by regulating movement of Rab11A-positive endosomes
Abstract
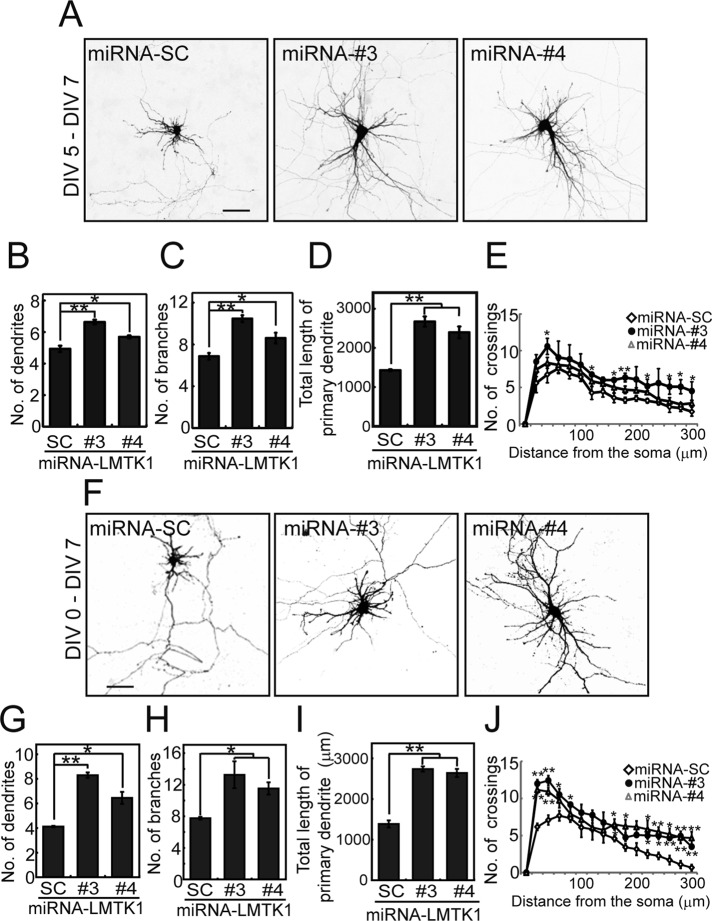
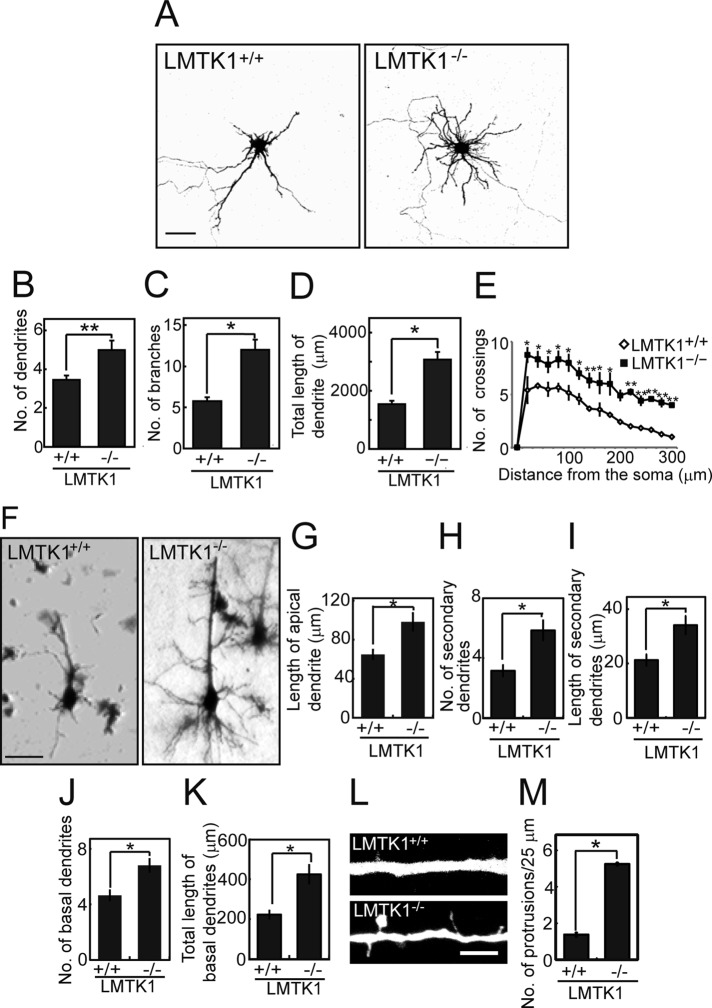
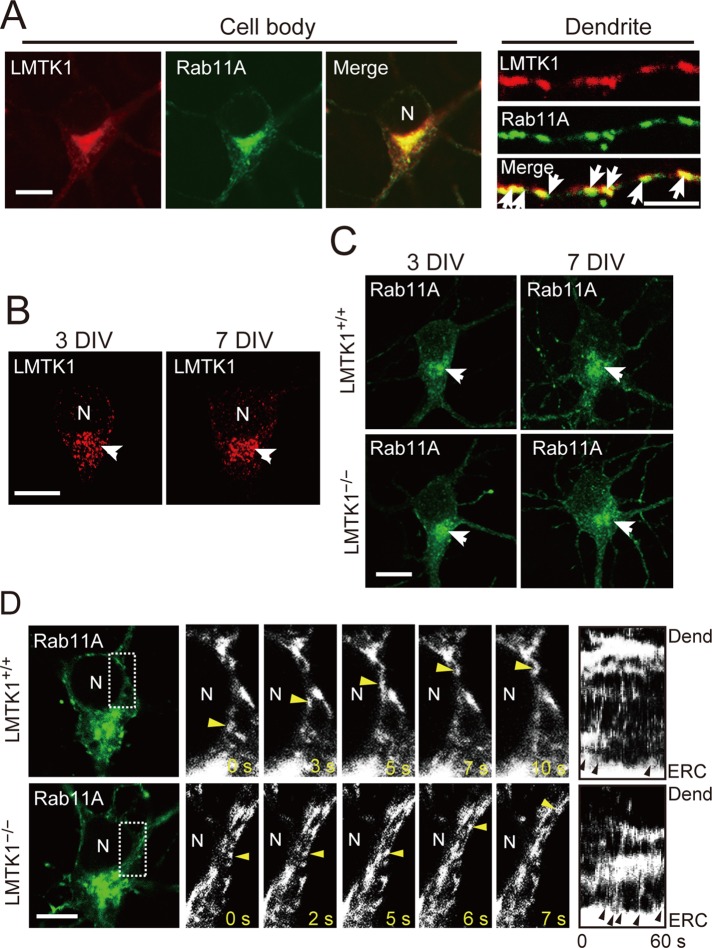
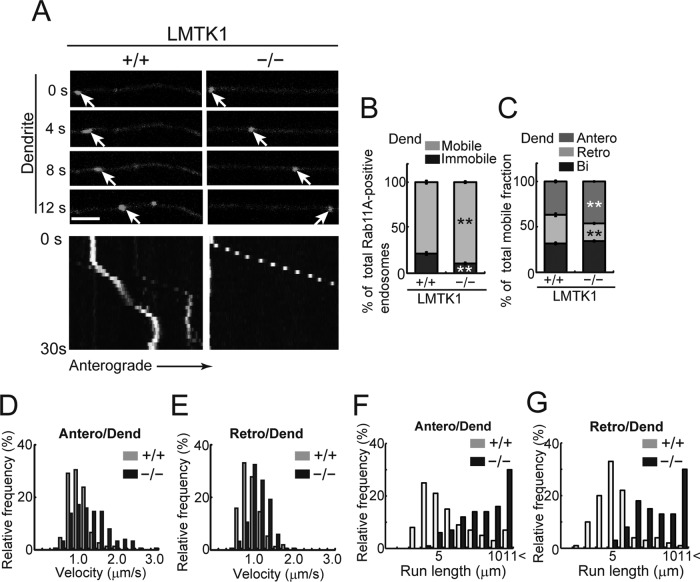
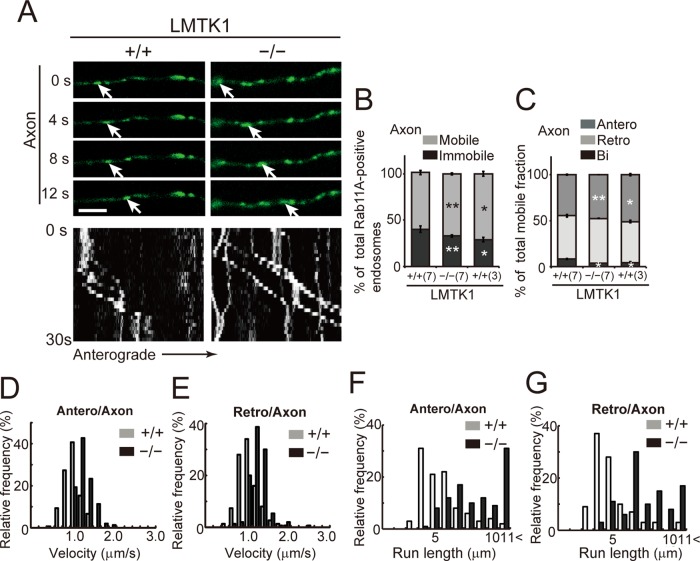
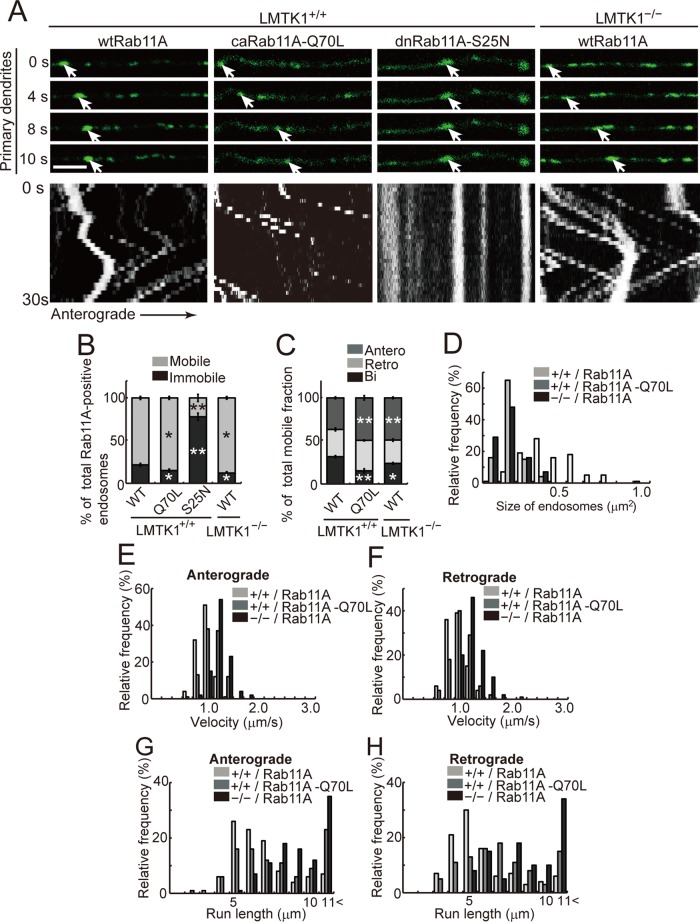
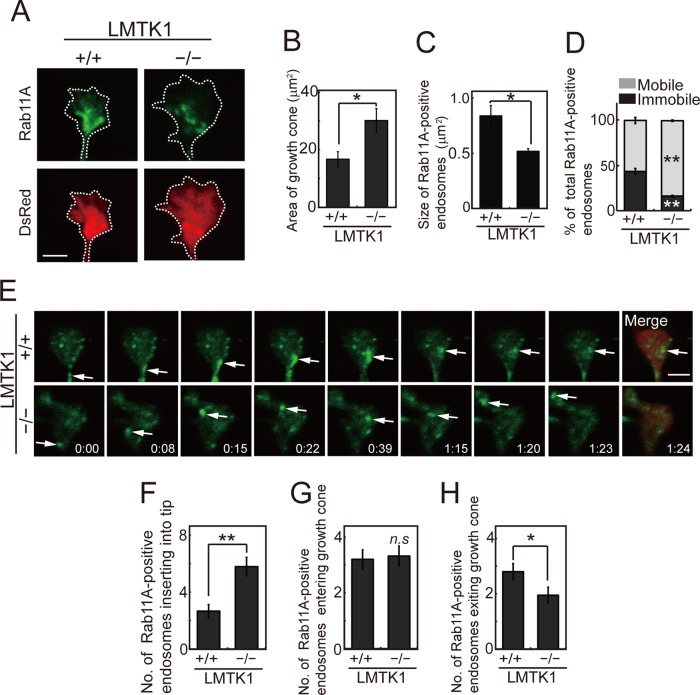
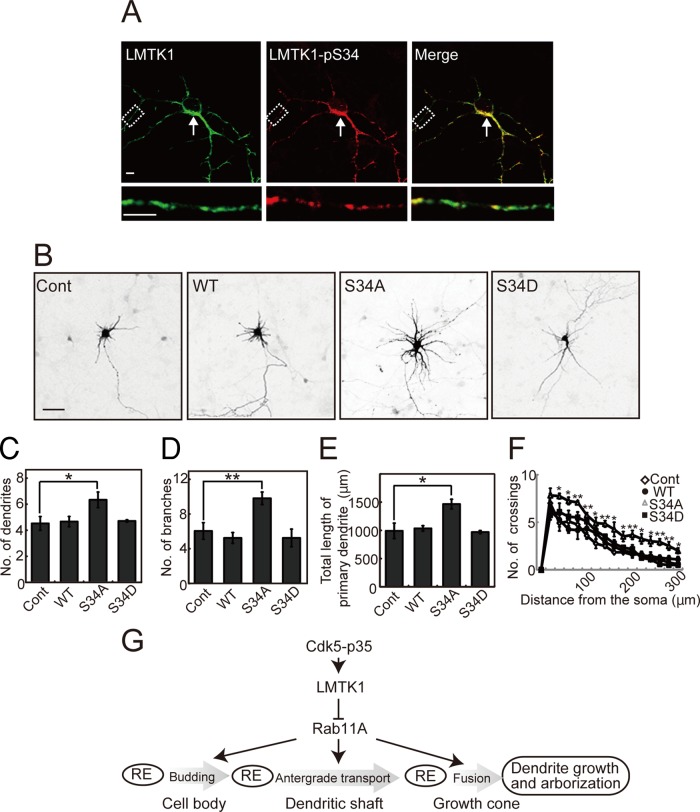
Neurons extend two types of neurites-axons and dendrites-that differ in structure and function. Although it is well understood that the cytoskeleton plays a pivotal role in neurite differentiation and extension, the mechanisms by which membrane components are supplied to growing axons or dendrites is largely unknown. We previously reported that the membrane supply to axons is regulated by lemur kinase 1 (LMTK1) through Rab11A-positive endosomes. Here we investigate the role of LMTK1 in dendrite formation. Down-regulation of LMTK1 increases dendrite growth and branching of cerebral cortical neurons in vitro and in vivo. LMTK1 knockout significantly enhances the prevalence, velocity, and run length of anterograde movement of Rab11A-positive endosomes to levels similar to those expressing constitutively active Rab11A-Q70L. Rab11A-positive endosome dynamics also increases in the cell body and growth cone of LMTK1-deficient neurons. Moreover, a nonphosphorylatable LMTK1 mutant (Ser34Ala, a Cdk5 phosphorylation site) dramatically promotes dendrite growth. Thus LMTK1 negatively controls dendritic formation by regulating Rab11A-positive endosomal trafficking in a Cdk5-dependent manner, indicating the Cdk5-LMTK1-Rab11A pathway as a regulatory mechanism of dendrite development as well as axon outgrowth.
© 2014 Takano et al. This article is distributed by The American Society for Cell Biology under license from the author(s). Two months after publication it is available to the public under an Attribution–Noncommercial–Share Alike 3.0 Unported Creative Commons License (http://creativecommons.org/licenses/by-nc-sa/3.0).
Figures
References
-
- Arimura N, Kaibuchi K. Neuronal polarity: from extracellular signals to intracellular mechanisms. Nat Rev Neurosci. 2007;8:194–205. - PubMed
-
- Baker SJ, Sumerson R, Reddy CD, Berrebi AS, Flynn DC, Reddy EP. Characterization of an alternatively spliced AATYK mRNA: expression pattern of AATYK in the brain and neuronal cells. Oncogene. 2001;20:1015–1021. - PubMed
-
- Bellon A. New genes associated with schizophrenia in neurite formation: a review of cell culture experiments. Mol Psychiatry. 2007;12:620–629. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
