

Operant conditioning of spinal reflexes: from basic science to clinical therapy
- PMID: 24672441
- PMCID: PMC3957063
- DOI: 10.3389/fnint.2014.00025
Operant conditioning of spinal reflexes: from basic science to clinical therapy
Abstract
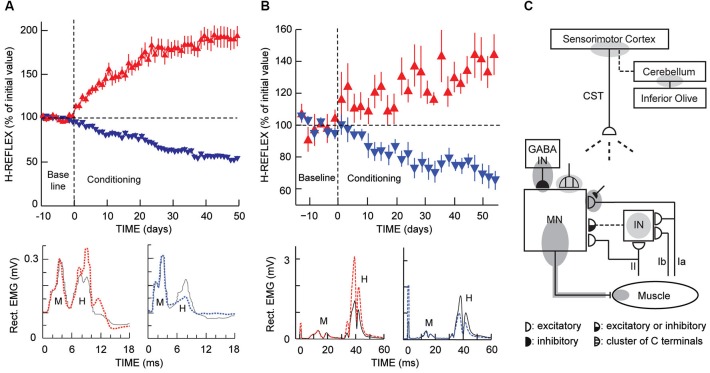
New appreciation of the adaptive capabilities of the nervous system, recent recognition that most spinal cord injuries are incomplete, and progress in enabling regeneration are generating growing interest in novel rehabilitation therapies. Here we review the 35-year evolution of one promising new approach, operant conditioning of spinal reflexes. This work began in the late 1970's as basic science; its purpose was to develop and exploit a uniquely accessible model for studying the acquisition and maintenance of a simple behavior in the mammalian central nervous system (CNS). The model was developed first in monkeys and then in rats, mice, and humans. Studies with it showed that the ostensibly simple behavior (i.e., a larger or smaller reflex) rests on a complex hierarchy of brain and spinal cord plasticity; and current investigations are delineating this plasticity and its interactions with the plasticity that supports other behaviors. In the last decade, the possible therapeutic uses of reflex conditioning have come under study, first in rats and then in humans. The initial results are very exciting, and they are spurring further studies. At the same time, the original basic science purpose and the new clinical purpose are enabling and illuminating each other in unexpected ways. The long course and current state of this work illustrate the practical importance of basic research and the valuable synergy that can develop between basic science questions and clinical needs.
Keywords: H-reflex; learning and memory; locomotion; spinal cord injury; spinal cord plasticity.
Figures

References
-
- Abel B. M., Emore E., Thompson A. K. (2011). Operant up-conditioning of the ankle dorsiflexor motor evoked potential in people with multiple sclerosis. Society for Neuroscience 41st Annual Meeting, Program No. 917.919. Washington, D.C.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical