

Mcm10 deficiency causes defective-replisome-induced mutagenesis and a dependency on error-free postreplicative repair
- PMID: 24674891
- PMCID: PMC4111720
- DOI: 10.4161/cc.28652
Mcm10 deficiency causes defective-replisome-induced mutagenesis and a dependency on error-free postreplicative repair
Abstract
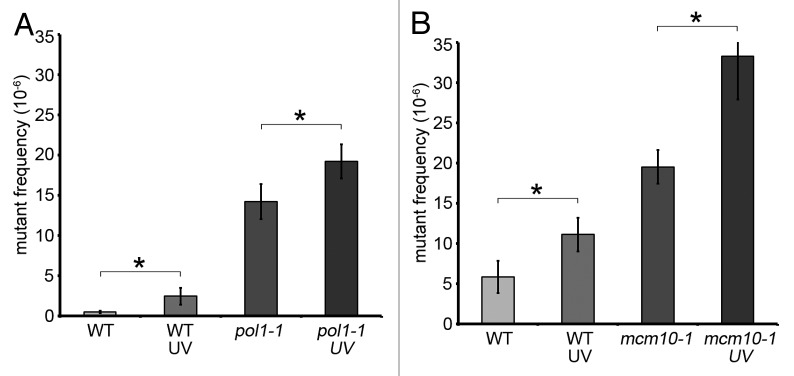
Mcm10 is a multifunctional replication factor with reported roles in origin activation, polymerase loading, and replication fork progression. The literature supporting these variable roles is controversial, and it has been debated whether Mcm10 has an active role in elongation. Here, we provide evidence that the mcm10-1 allele confers alterations in DNA synthesis that lead to defective-replisome-induced mutagenesis (DRIM). Specifically, we observed that mcm10-1 cells exhibited elevated levels of PCNA ubiquitination and activation of the translesion polymerase, pol-ζ. Whereas translesion synthesis had no measurable impact on viability, mcm10-1 mutants also engaged in error-free postreplicative repair (PRR), and this pathway promoted survival at semi-permissive conditions. Replication gaps in mcm10-1 were likely caused by elongation defects, as dbf4-1 mutants, which are compromised for origin activation did not display any hallmarks of replication stress. Furthermore, we demonstrate that deficiencies in priming, induced by a pol1-1 mutation, also resulted in DRIM, but not in error-free PRR. Similar to mcm10-1 mutants, DRIM did not rescue the replication defect in pol1-1 cells. Thus, it appears that DRIM is not proficient to fill replication gaps in pol1-1 and mcm10-1 mutants. Moreover, the ability to correctly prime nascent DNA may be a crucial prerequisite to initiate error-free PRR.
Keywords: DNA replication; DRIM; Mcm10; PCNA ubiquitination; Pol1; sumoylation; translesion synthesis.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous