

Innate immune factors modulate ethanol interaction with GABAergic transmission in mouse central amygdala
- PMID: 24675033
- PMCID: PMC4126651
- DOI: 10.1016/j.bbi.2014.03.007
Innate immune factors modulate ethanol interaction with GABAergic transmission in mouse central amygdala
Abstract
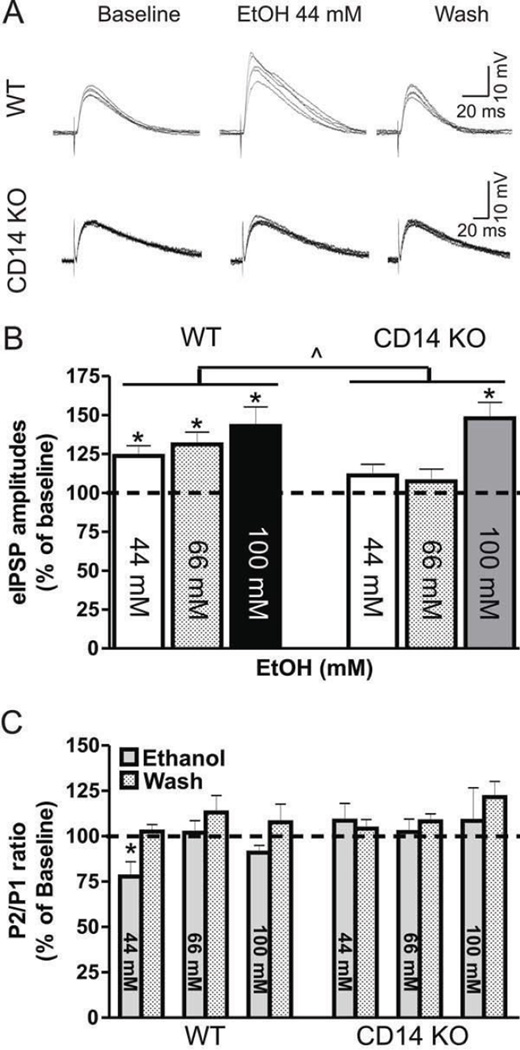
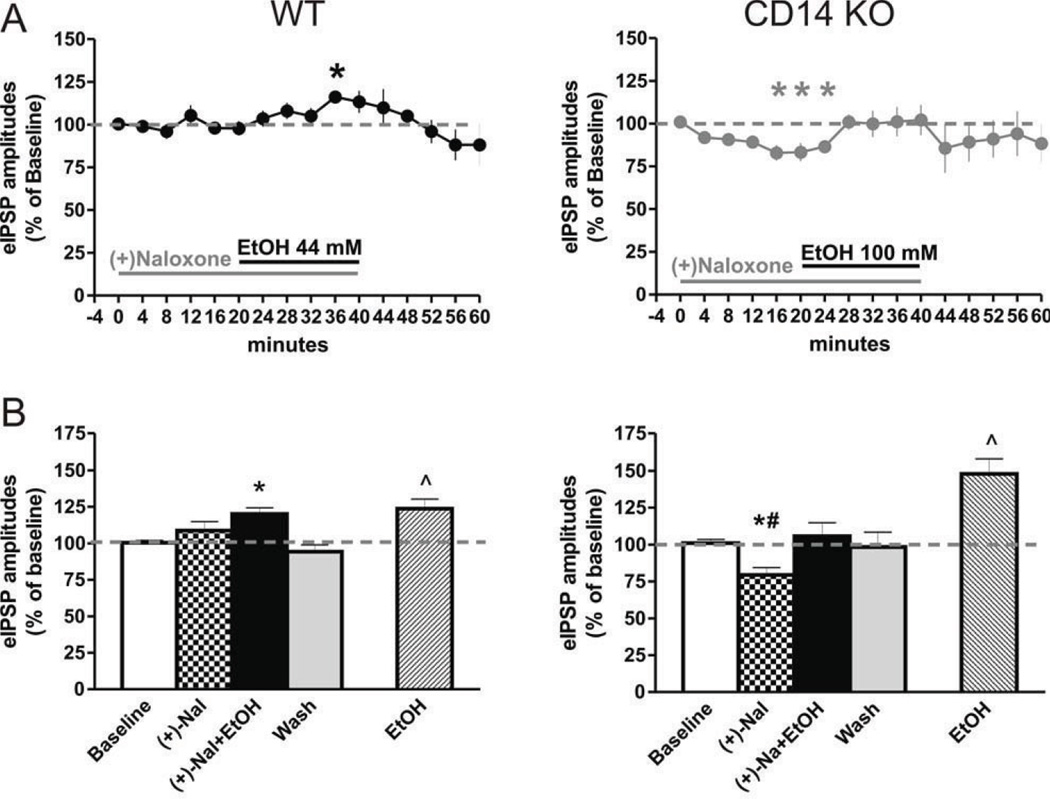
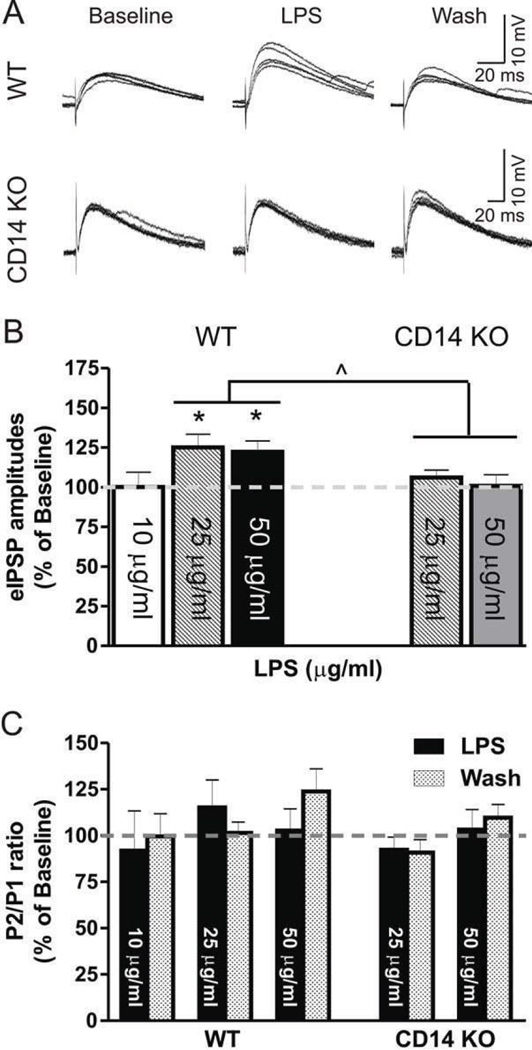
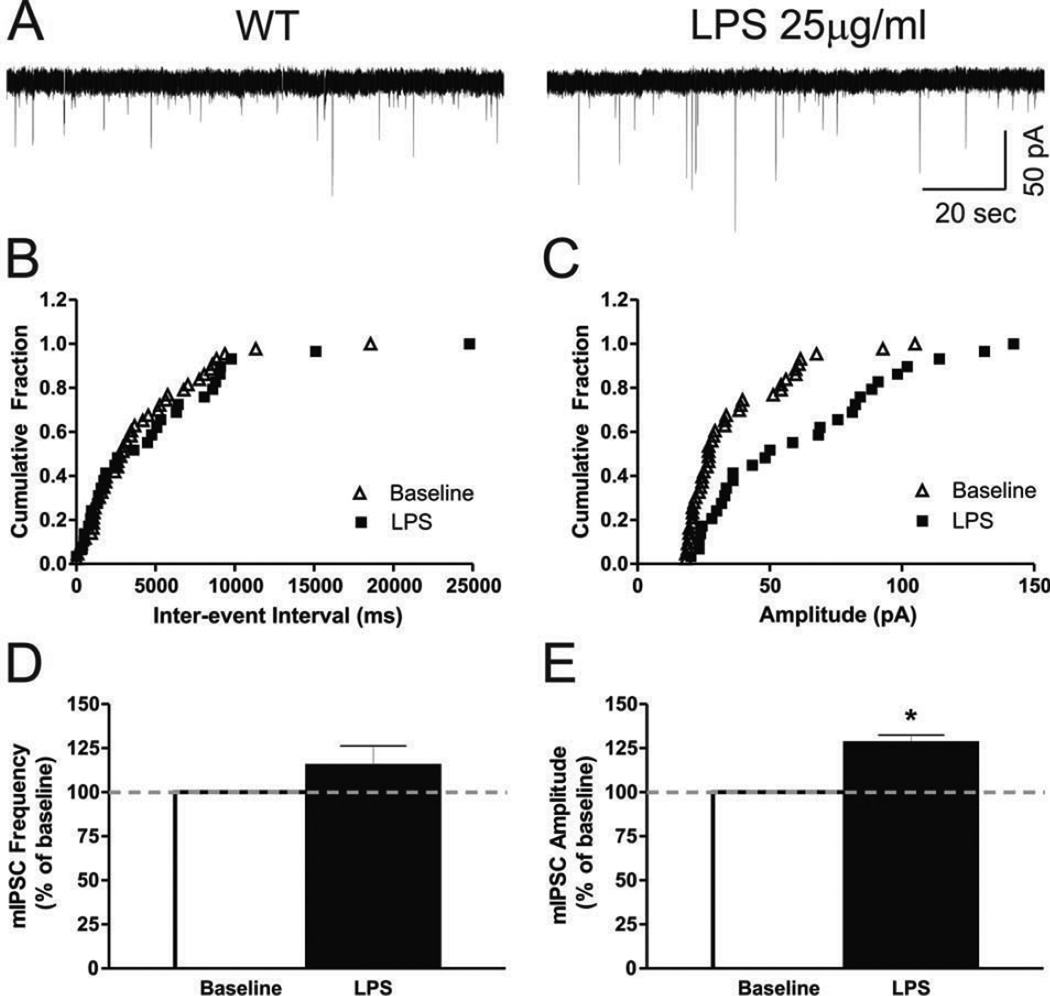
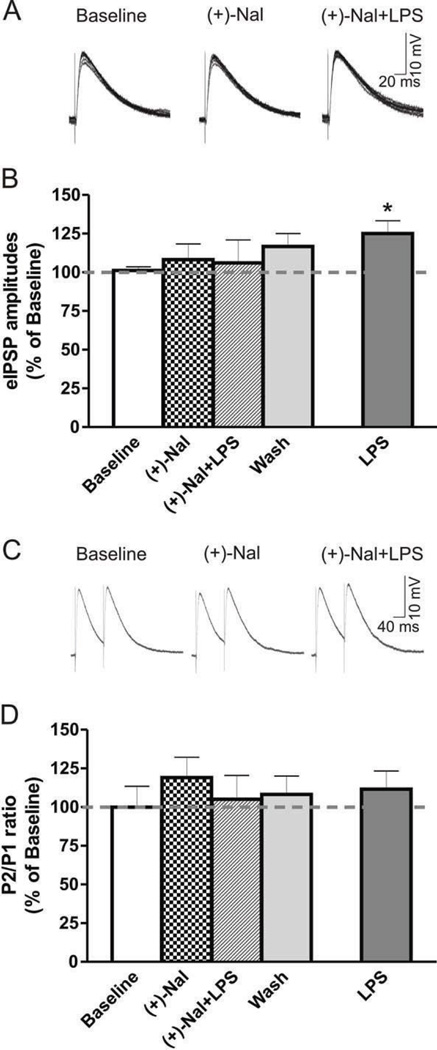
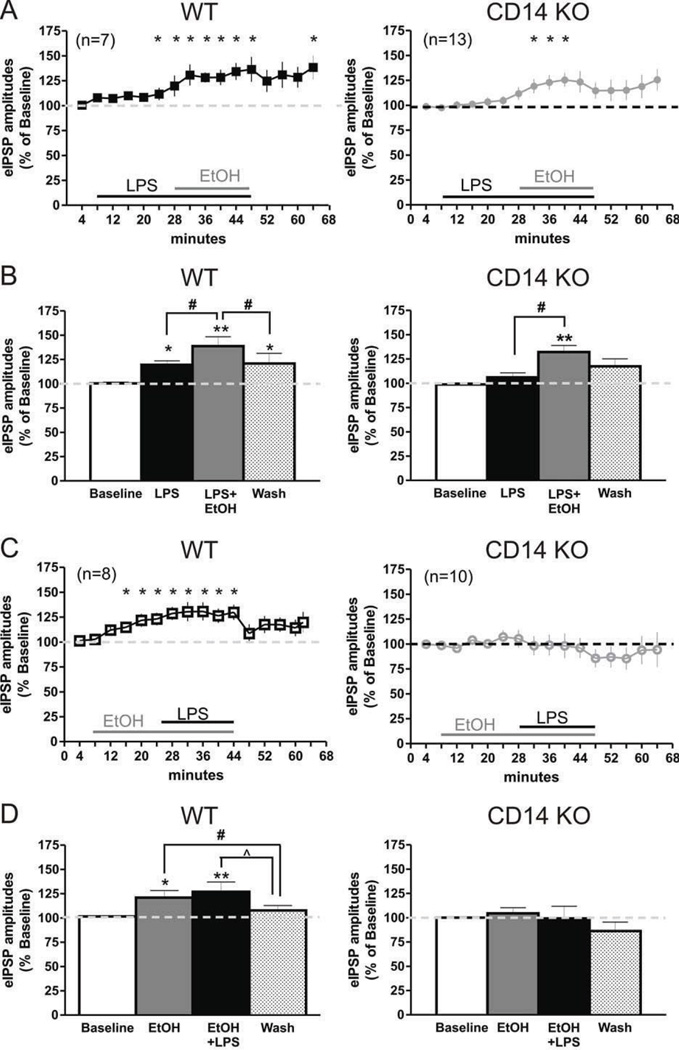
Excessive ethanol drinking in rodent models may involve activation of the innate immune system, especially toll-like receptor 4 (TLR4) signaling pathways. We used intracellular recording of evoked GABAergic inhibitory postsynaptic potentials (eIPSPs) in central amygdala (CeA) neurons to examine the role of TLR4 activation by lipopolysaccharide (LPS) and deletion of its adapter protein CD14 in acute ethanol effects on the GABAergic system. Ethanol (44, 66 or 100mM) and LPS (25 and 50μg/ml) both augmented eIPSPs in CeA of wild type (WT) mice. Ethanol (44mM) decreased paired-pulse facilitation (PPF), suggesting a presynaptic mechanism of action. Acute LPS (25μg/ml) had no effect on PPF and significantly increased the mean miniature IPSC amplitude, indicating a postsynaptic mechanism of action. Acute LPS pre-treatment potentiated ethanol (44mM) effects on eIPSPs in WT mice and restored ethanol's augmenting effects on the eIPSP amplitude in CD14 knockout (CD14 KO) mice. Both the LPS and ethanol (44-66mM) augmentation of eIPSPs was diminished significantly in most CeA neurons of CD14 KO mice; however, ethanol at the highest concentration tested (100mM) still increased eIPSP amplitudes. By contrast, ethanol pre-treatment occluded LPS augmentation of eIPSPs in WT mice and had no significant effect in CD14 KO mice. Furthermore, (+)-naloxone, a TLR4-MD-2 complex inhibitor, blocked LPS effects on eIPSPs in WT mice and delayed the ethanol-induced potentiation of GABAergic transmission. In CeA neurons of CD14 KO mice, (+)-naloxone alone diminished eIPSPs, and subsequent co-application of 100mM ethanol restored the eIPSPs to baseline levels. In summary, our results indicate that TLR4 and CD14 signaling play an important role in the acute ethanol effects on GABAergic transmission in the CeA and support the idea that CD14 and TLR4 may be therapeutic targets for treatment of alcohol abuse.
Keywords: Alcohol; Drug abuse; Extended amygdala; Inflammation; LPS; Neuroimmune; Toll-like receptor.
Copyright © 2014 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Acosta C, Davies A. Bacterial lipopolysaccharide regulates nociceptin expression in sensory neurons. J Neurosci Res. 2008;86:1077–1086. - PubMed
-
- Alfonso-Loeches S, Guerri C. Molecular and behavioral aspects of the actions of alcohol on the adult and developing brain. Crit Rev Clin Lab Sci. 2011;48:19–47. - PubMed
-
- Alfonso-Loeches S, Pascual M, Gomez-Pinedo U, Pascual-Lucas M, Renau-Piqueras J, Guerri C. Toll-like receptor 4 participates in the myelin disruptions associated with chronic alcohol abuse. Glia. 2012;60:948–964. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
