

Investigation of inflammation and tissue patterning in the gut using a Spatially Explicit General-purpose Model of Enteric Tissue (SEGMEnT)
- PMID: 24675765
- PMCID: PMC3967920
- DOI: 10.1371/journal.pcbi.1003507
Investigation of inflammation and tissue patterning in the gut using a Spatially Explicit General-purpose Model of Enteric Tissue (SEGMEnT)
Abstract
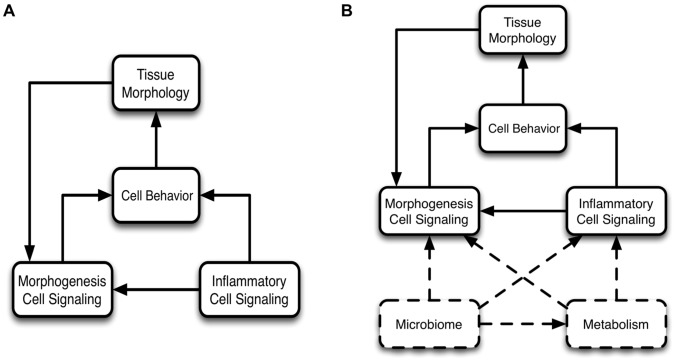
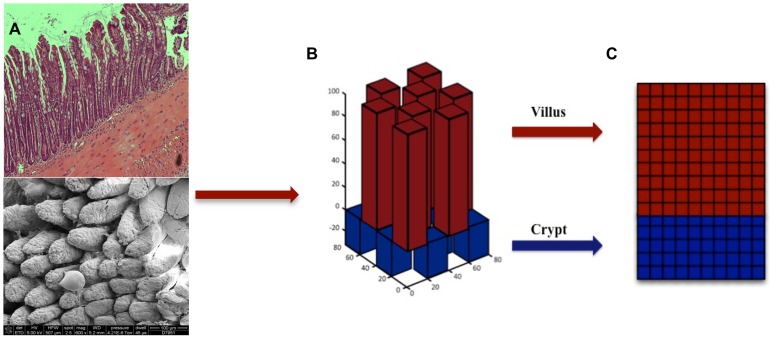
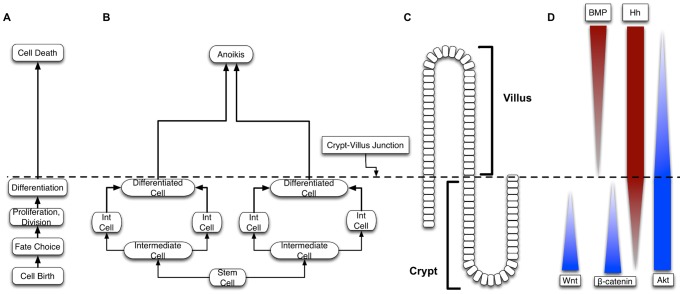
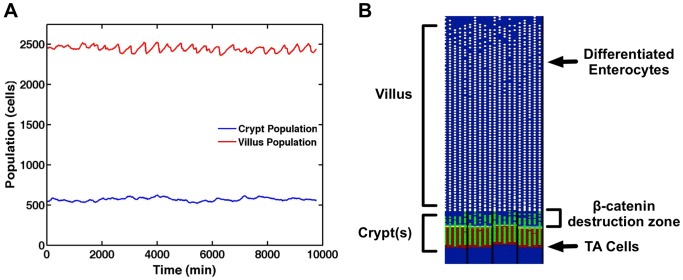
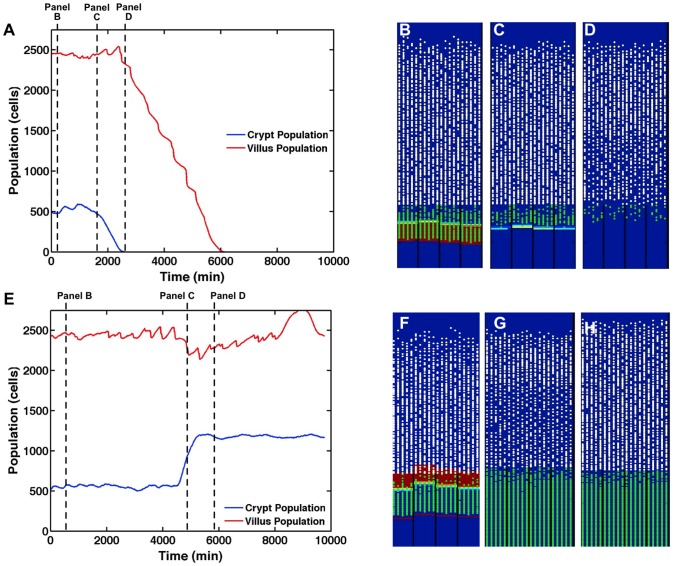
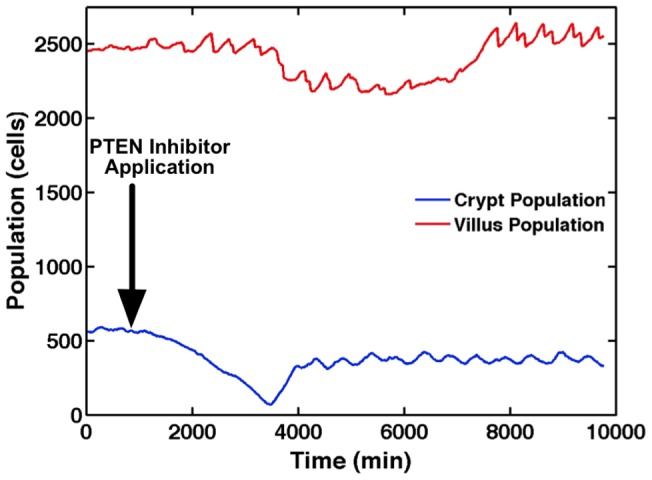
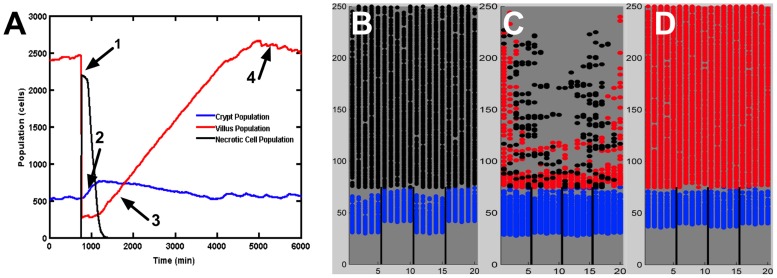
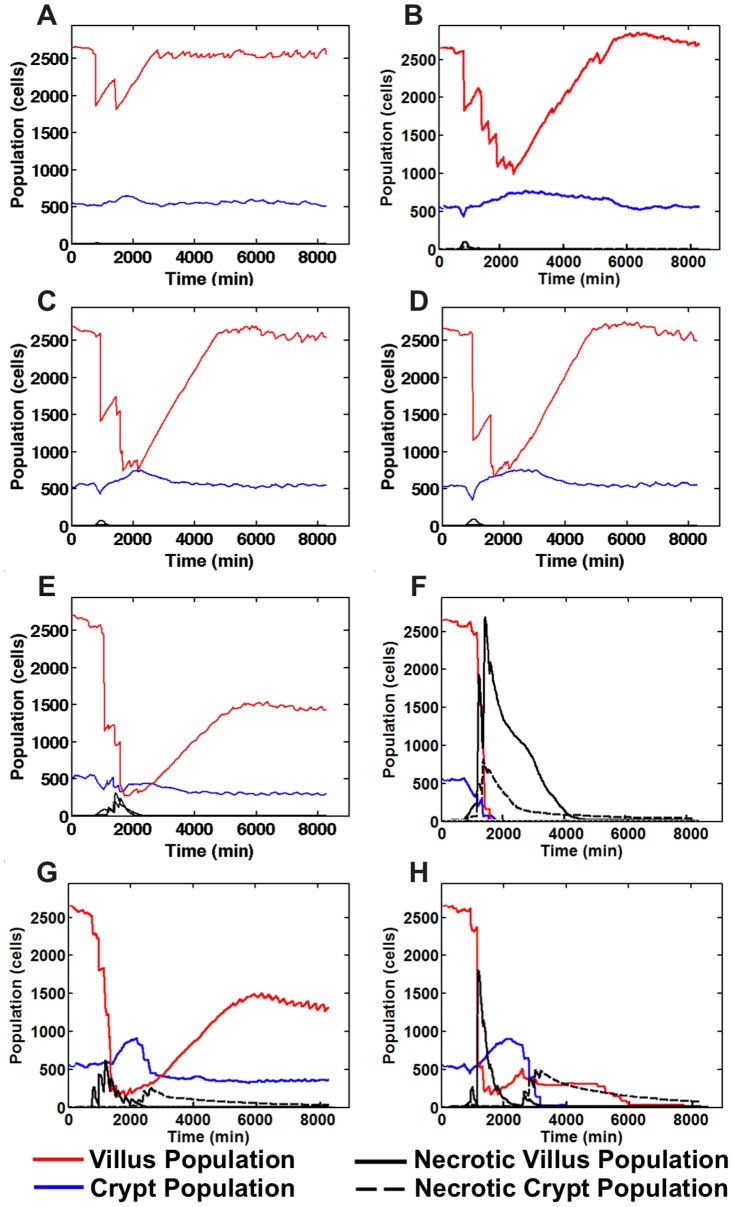
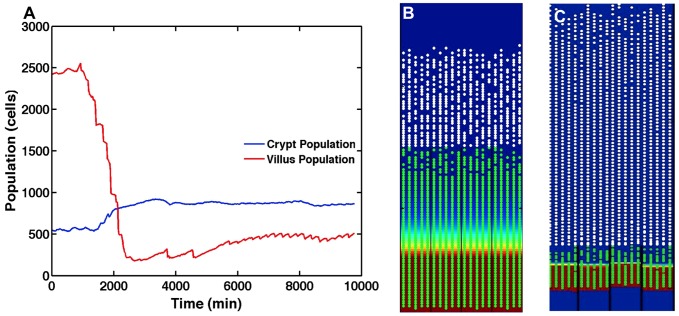
The mucosa of the intestinal tract represents a finely tuned system where tissue structure strongly influences, and is turn influenced by, its function as both an absorptive surface and a defensive barrier. Mucosal architecture and histology plays a key role in the diagnosis, characterization and pathophysiology of a host of gastrointestinal diseases. Inflammation is a significant factor in the pathogenesis in many gastrointestinal diseases, and is perhaps the most clinically significant control factor governing the maintenance of the mucosal architecture by morphogenic pathways. We propose that appropriate characterization of the role of inflammation as a controller of enteric mucosal tissue patterning requires understanding the underlying cellular and molecular dynamics that determine the epithelial crypt-villus architecture across a range of conditions from health to disease. Towards this end we have developed the Spatially Explicit General-purpose Model of Enteric Tissue (SEGMEnT) to dynamically represent existing knowledge of the behavior of enteric epithelial tissue as influenced by inflammation with the ability to generate a variety of pathophysiological processes within a common platform and from a common knowledge base. In addition to reproducing healthy ileal mucosal dynamics as well as a series of morphogen knock-out/inhibition experiments, SEGMEnT provides insight into a range of clinically relevant cellular-molecular mechanisms, such as a putative role for Phosphotase and tensin homolog/phosphoinositide 3-kinase (PTEN/PI3K) as a key point of crosstalk between inflammation and morphogenesis, the protective role of enterocyte sloughing in enteric ischemia-reperfusion and chronic low level inflammation as a driver for colonic metaplasia. These results suggest that SEGMEnT can serve as an integrating platform for the study of inflammation in gastrointestinal disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Turner JR (2004) The Gastrointestinal Tract. In: Vinay Kumar AKA, Jon C. Aster, Nelson Fausto, Robbins and Cotran: Pathological Basis of Disease, 8th Edition. Philadelphia, PA: Saunders-Elsevier.
-
- Seal JB, Morowitz M, Zaborina O, An G, Alverdy JC (2010) The molecular Koch's postulates and surgical infection: a view forward. Surgery 147: 757–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
