

Parent-of-origin effects implicate epigenetic regulation of experimental autoimmune encephalomyelitis and identify imprinted Dlk1 as a novel risk gene
- PMID: 24676147
- PMCID: PMC3967983
- DOI: 10.1371/journal.pgen.1004265
Parent-of-origin effects implicate epigenetic regulation of experimental autoimmune encephalomyelitis and identify imprinted Dlk1 as a novel risk gene
Abstract
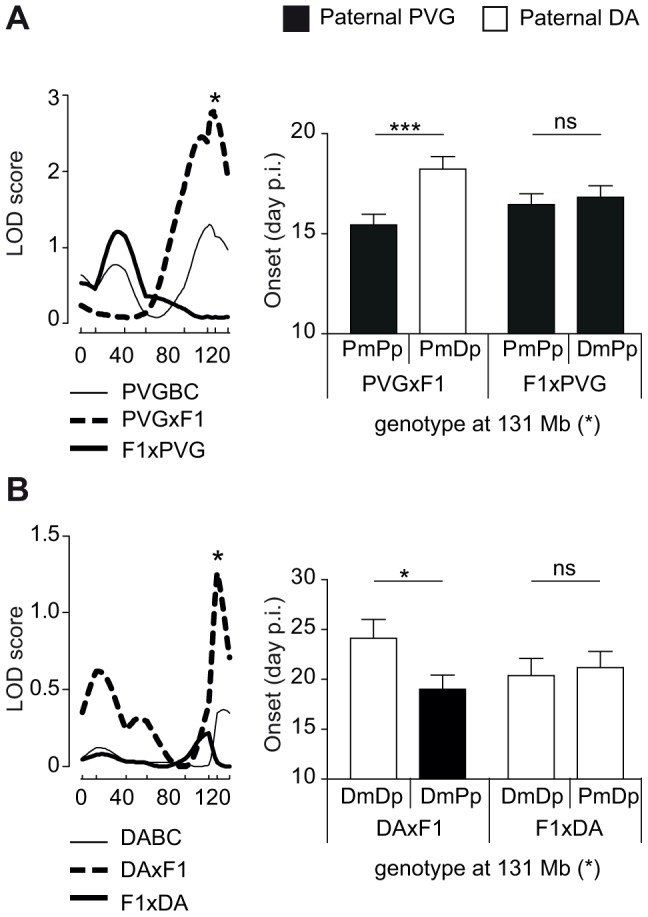
Parent-of-origin effects comprise a range of genetic and epigenetic mechanisms of inheritance. Recently, detection of such effects implicated epigenetic mechanisms in the etiology of multiple sclerosis (MS), a chronic inflammatory disease of the central nervous system. We here sought to dissect the magnitude and the type of parent-of-origin effects in the pathogenesis of experimental neuroinflammation under controlled environmental conditions. We investigated inheritance of an MS-like disease in rat, experimental autoimmune encephalomyelitis (EAE), using a backcross strategy designed to identify the parental origin of disease-predisposing alleles. A striking 37-54% of all detected disease-predisposing loci depended on parental transmission. Additionally, the Y chromosome from the susceptible strain contributed to disease susceptibility. Accounting for parent-of-origin enabled more powerful and precise identification of novel risk factors and increased the disease variance explained by the identified factors by 2-4-fold. The majority of loci displayed an imprinting-like pattern whereby a gene expressed only from the maternal or paternal copy exerts an effect. In particular, a locus on chromosome 6 comprises a well-known cluster of imprinted genes including the paternally expressed Dlk1, an atypical Notch ligand. Disease-predisposing alleles at the locus conferred lower Dlk1 expression in rats and, together with data from transgenic overexpressing Dlk1 mice, demonstrate that reduced Dlk1 drives more severe disease and modulates adaptive immune reactions in EAE. Our findings suggest a significant epigenetic contribution to the etiology of EAE. Incorporating these effects enables more powerful and precise identification of novel risk factors with diagnostic and prognostic implications for complex disease.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
