

Evolutionary convergence of cell-specific gene expression in independent lineages of C4 grasses
- PMID: 24676859
- PMCID: PMC4012605
- DOI: 10.1104/pp.114.238667
Evolutionary convergence of cell-specific gene expression in independent lineages of C4 grasses
Abstract
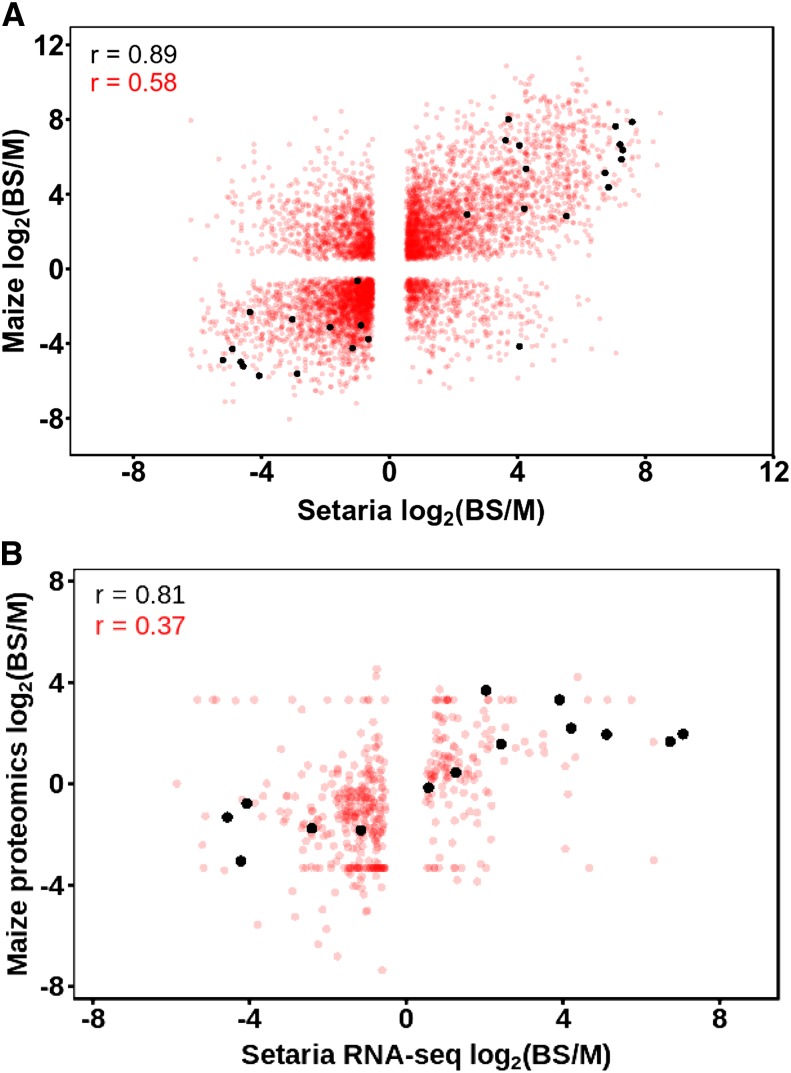
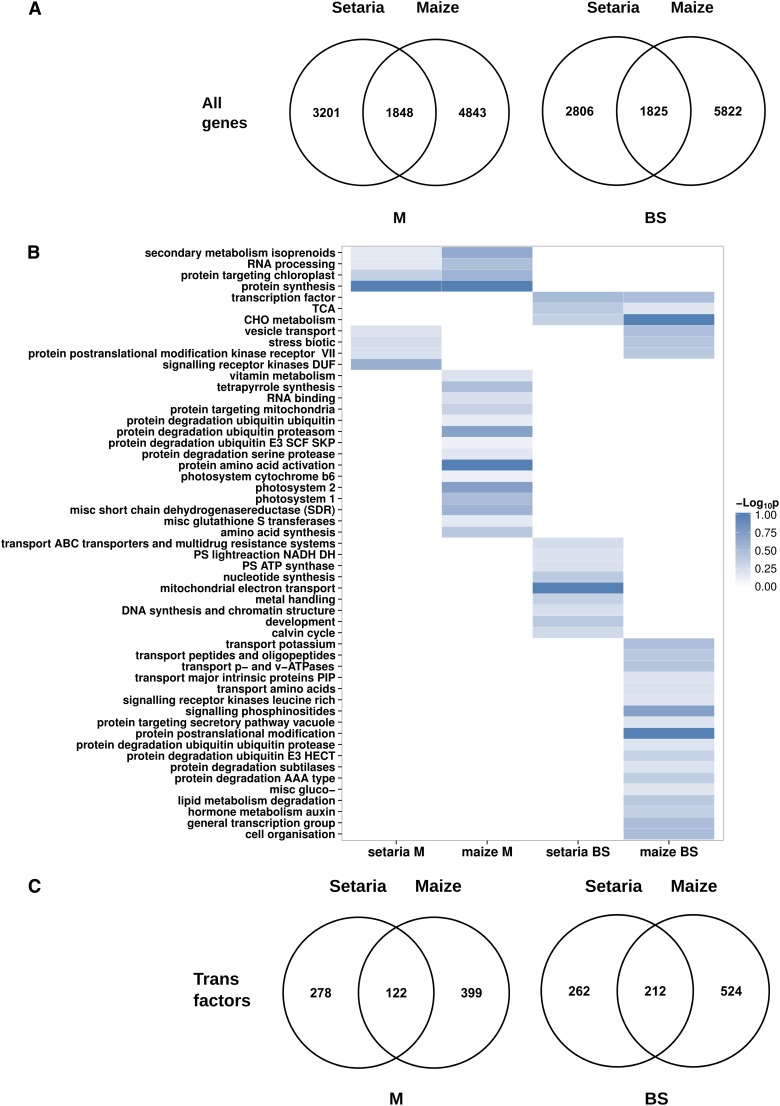
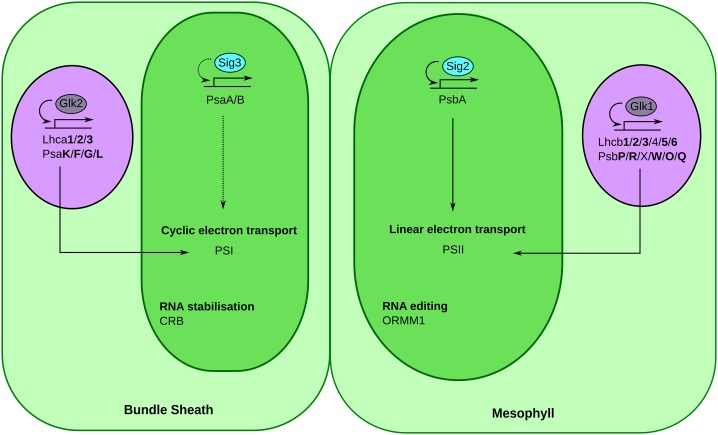
Leaves of almost all C4 lineages separate the reactions of photosynthesis into the mesophyll (M) and bundle sheath (BS). The extent to which messenger RNA profiles of M and BS cells from independent C4 lineages resemble each other is not known. To address this, we conducted deep sequencing of RNA isolated from the M and BS of Setaria viridis and compared these data with publicly available information from maize (Zea mays). This revealed a high correlation (r=0.89) between the relative abundance of transcripts encoding proteins of the core C4 pathway in M and BS cells in these species, indicating significant convergence in transcript accumulation in these evolutionarily independent C4 lineages. We also found that the vast majority of genes encoding proteins of the C4 cycle in S. viridis are syntenic to homologs used by maize. In both lineages, 122 and 212 homologous transcription factors were preferentially expressed in the M and BS, respectively. Sixteen shared regulators of chloroplast biogenesis were identified, 14 of which were syntenic homologs in maize and S. viridis. In sorghum (Sorghum bicolor), a third C4 grass, we found that 82% of these trans-factors were also differentially expressed in either M or BS cells. Taken together, these data provide, to our knowledge, the first quantification of convergence in transcript abundance in the M and BS cells from independent lineages of C4 grasses. Furthermore, the repeated recruitment of syntenic homologs from large gene families strongly implies that parallel evolution of both structural genes and trans-factors underpins the polyphyletic evolution of this highly complex trait in the monocotyledons.
Figures
References
-
- Anders S (2011) HTSeq: analysing high-throughput sequencing data with Python. http://www-huber.embl.de/users/anders/HTSeq/ (April 7, 2014) - PMC - PubMed
-
- Aubry S, Brown NJ, Hibberd JM. (2011) The role of proteins in C3 plants prior to their recruitment into the C4 pathway. J Exp Bot 62: 3049–3059 - PubMed
-
- Bennetzen JL, Schmutz J, Wang H, Percifield R, Hawkins J, Pontaroli AC, Estep M, Feng L, Vaughn JN, Grimwood J, et al. (2012) Reference genome sequence of the model plant Setaria. Nat Biotechnol 30: 555–561 - PubMed
-
- Brown NJ, Newell CA, Stanley S, Chen JE, Perrin AJ, Kajala K, Hibberd JM. (2011) Independent and parallel recruitment of preexisting mechanisms underlying C4 photosynthesis. Science 331: 1436–1439 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
