

Tendon-to-bone attachment: from development to maturity
- PMID: 24677726
- PMCID: PMC4076491
- DOI: 10.1002/bdrc.21056
Tendon-to-bone attachment: from development to maturity
Abstract
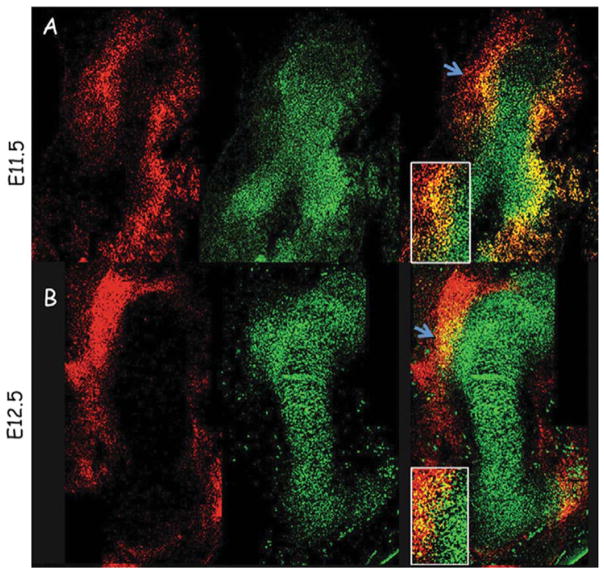
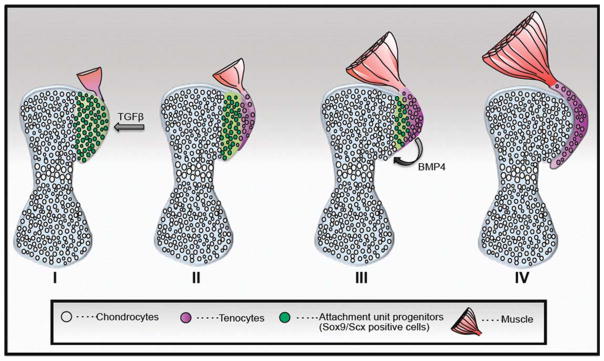
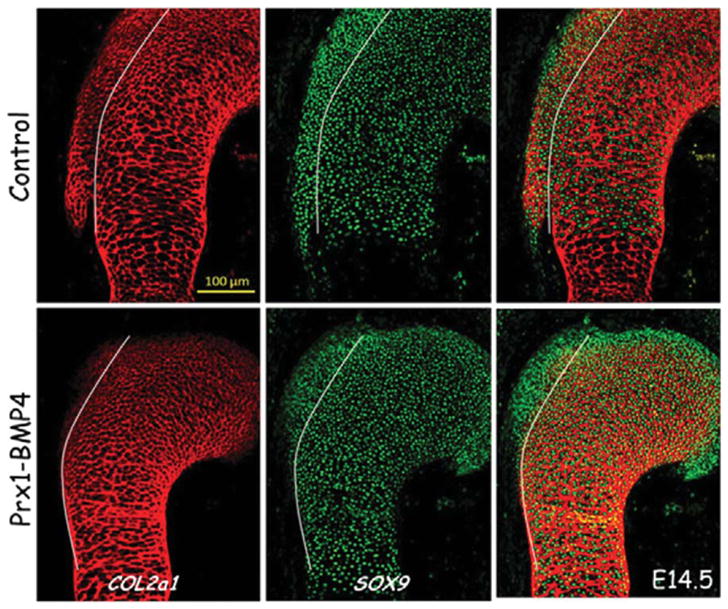
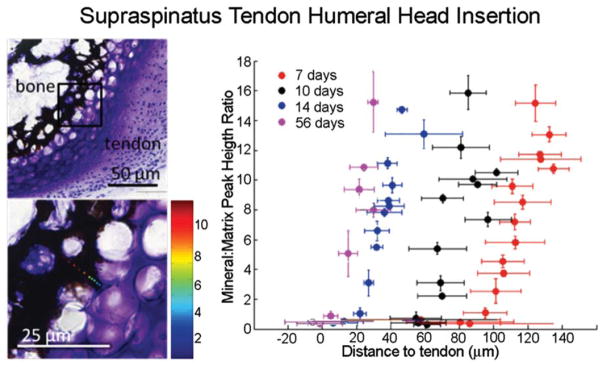
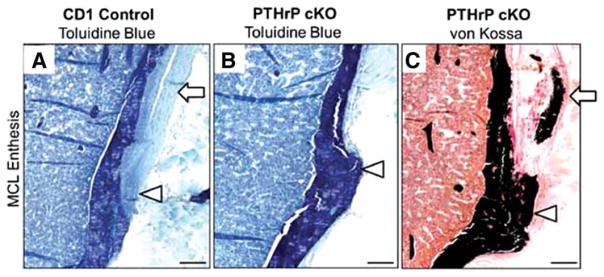
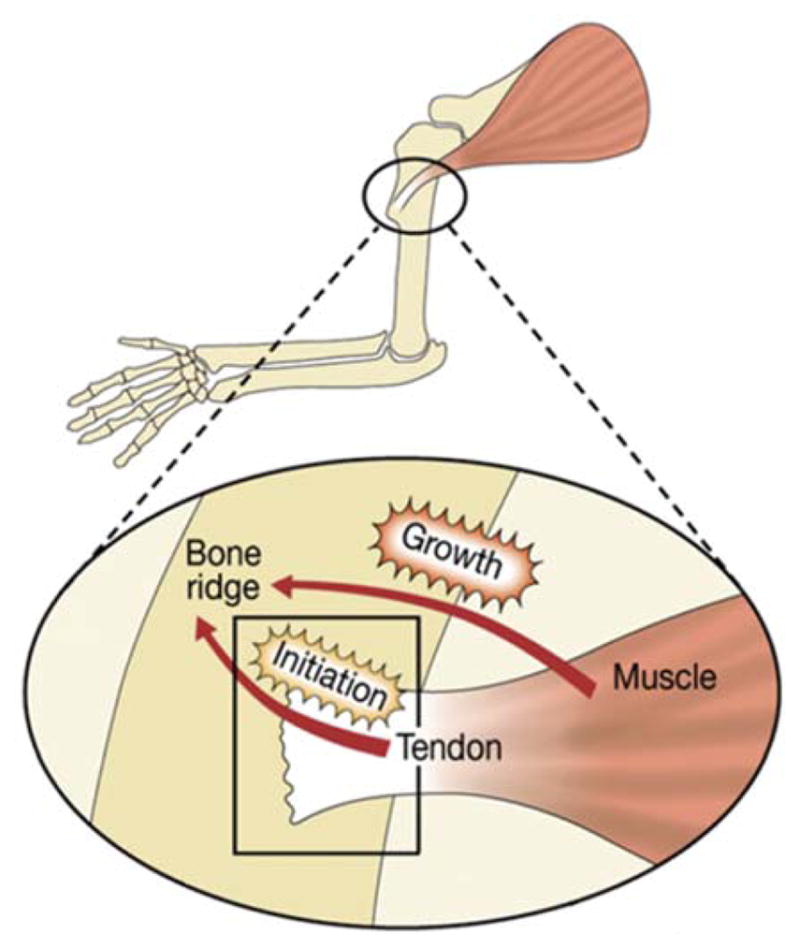
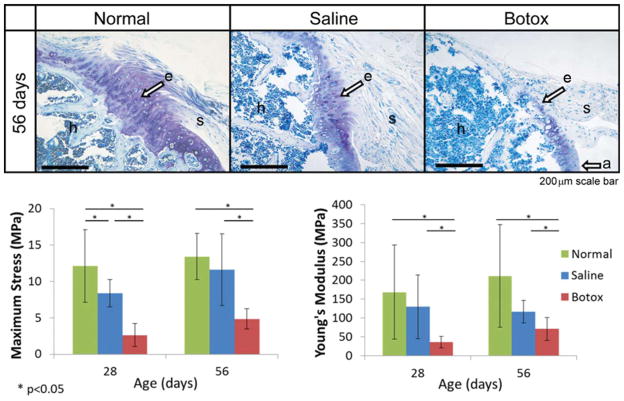
The attachment between tendon and bone occurs across a complex transitional tissue that minimizes stress concentrations and allows for load transfer between muscles and skeleton. This unique tissue cannot be reconstructed following injury, leading to high incidence of recurrent failure and stressing the need for new clinical approaches. This review describes the current understanding of the development and function of the attachment site between tendon and bone. The embryonic attachment unit, namely, the tip of the tendon and the bone eminence into which it is inserted, was recently shown to develop modularly from a unique population of Sox9- and Scx-positive cells, which are distinct from tendon fibroblasts and chondrocytes. The fate and differentiation of these cells is regulated by transforming growth factor beta and bone morphogenetic protein signaling, respectively. Muscle loads are then necessary for the tissue to mature and mineralize. Mineralization of the attachment unit, which occurs postnatally at most sites, is largely controlled by an Indian hedgehog/parathyroid hormone-related protein feedback loop. A number of fundamental questions regarding the development of this remarkable attachment system require further study. These relate to the signaling mechanism that facilitates the formation of an interface with a gradient of cellular and extracellular phenotypes, as well as to the interactions between tendon and bone at the point of attachment.
Keywords: BMP4; IHH; PTHrP; SCX; bone; bone eminence; cartilage; enthesis; musculoskeletal development; tendon insertion.
Copyright © 2014 Wiley Periodicals, Inc.
Conflict of interest statement
The authors have no conflicts of interest with the material reviewed in this article.
Figures
References
-
- Benjamin M, Kumai T, Milz S, Boszczyk BM, et al. The skeletal attachment of tendons—tendon “enthuses”. Comp Biochem Physiol A Mol Integr Physiol. 2002;133:931–945. - PubMed
-
- Biewener AA, Fazzalari NL, Konieczynski DD, Baudinette RV. Adaptive changes in trabecular architecture in relation to functional strain patterns and disuse. Bone. 1996;19:1–8. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
