

Differences in the composition of the human antibody repertoire by B cell subsets in the blood
- PMID: 24678310
- PMCID: PMC3958703
- DOI: 10.3389/fimmu.2014.00096
Differences in the composition of the human antibody repertoire by B cell subsets in the blood
Abstract
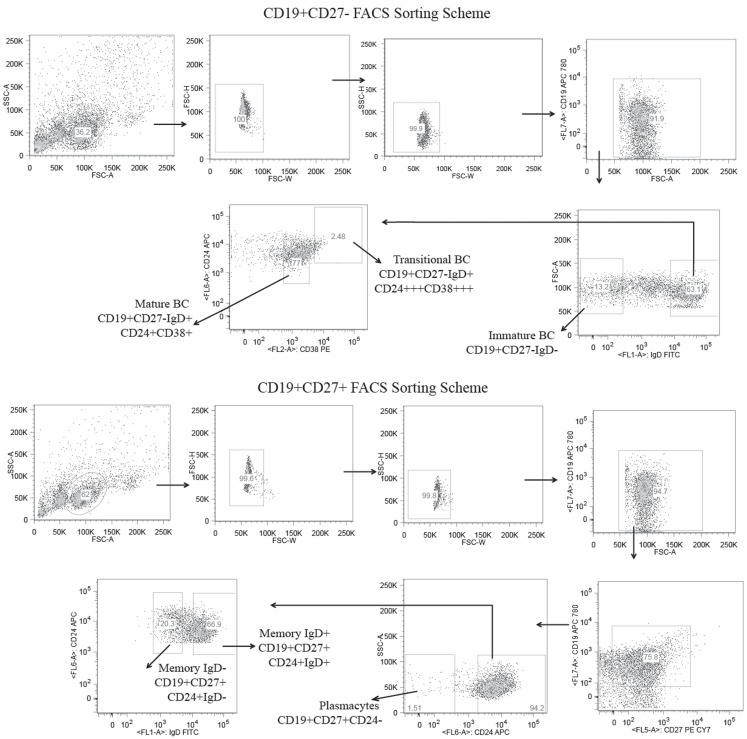
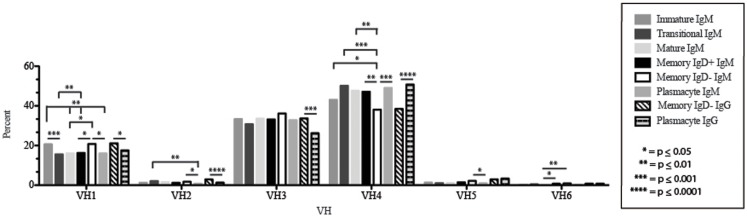
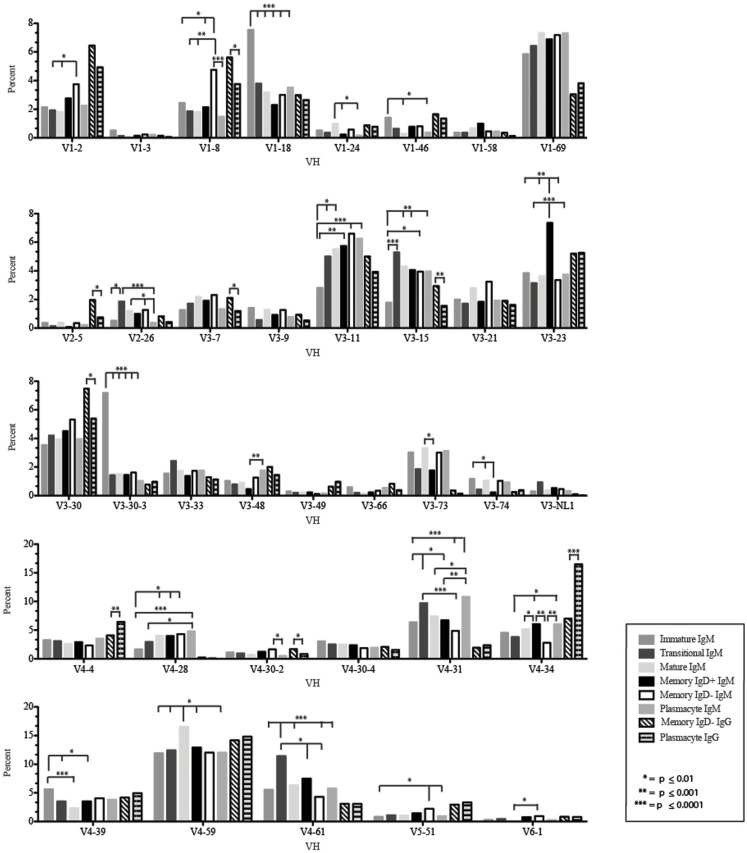
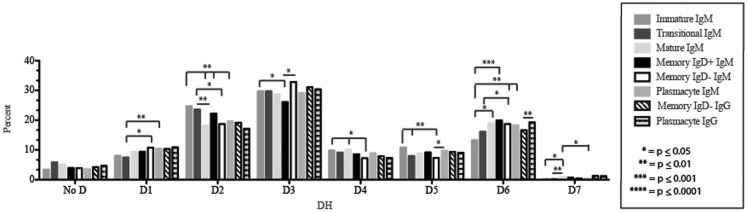
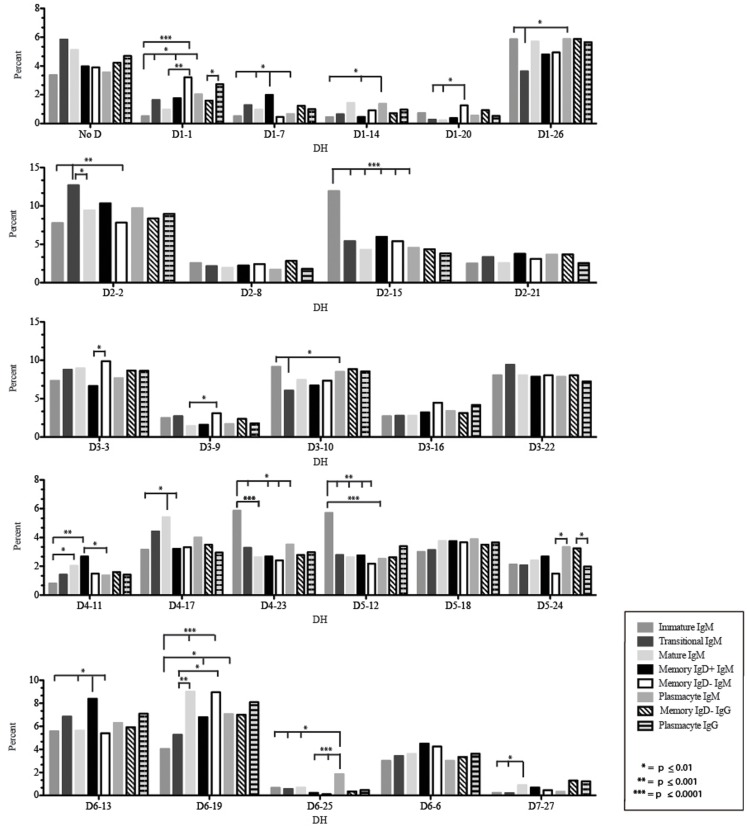
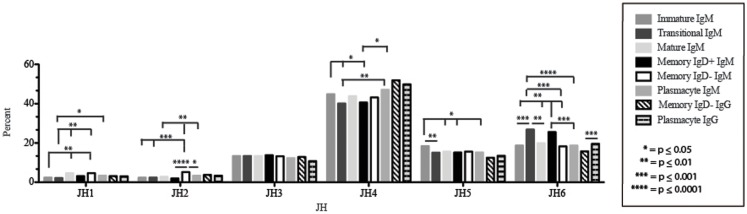
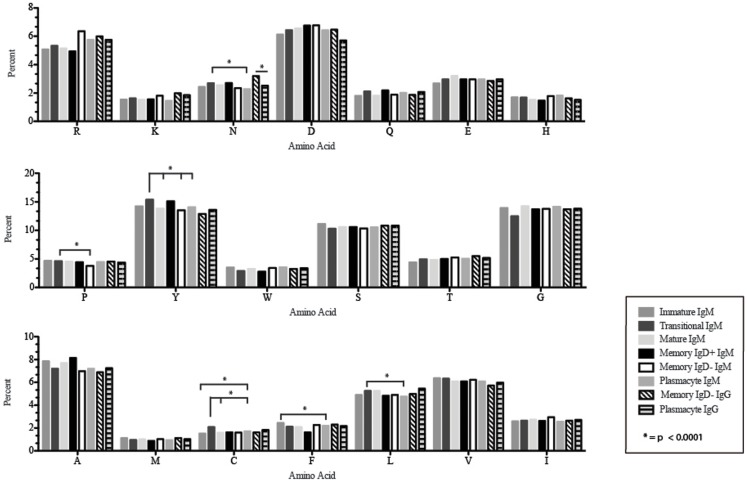
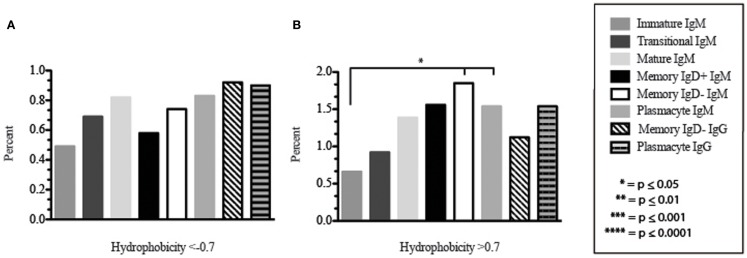
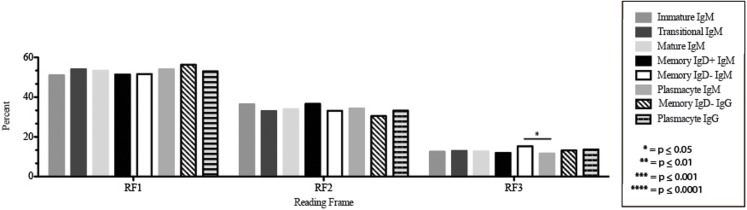
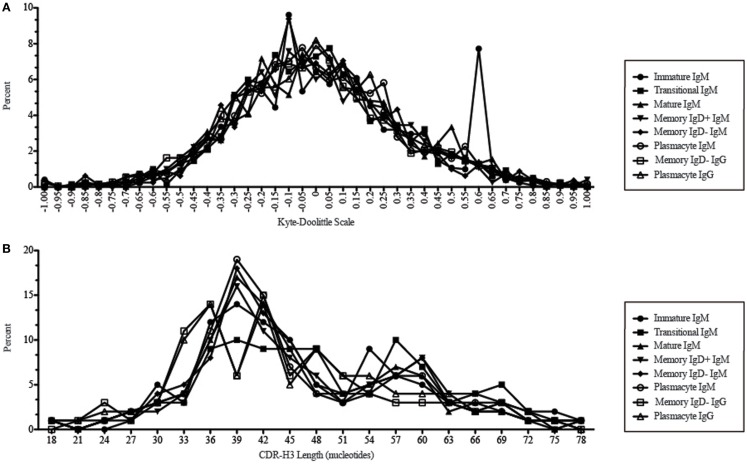
The vast initial diversity of the antibody repertoire is generated centrally by means of a complex series of V(D)J gene rearrangement events, variation in the site of gene segment joining, and TdT catalyzed N-region addition. Although the diversity is great, close inspection has revealed distinct and unique characteristics in the antibody repertoires expressed by different B cell developmental subsets. In order to illustrate our approach to repertoire analysis, we present an in-depth comparison of V(D)J gene usage, hydrophobicity, length, DH reading frame, and amino acid usage between heavy chain repertoires expressed by immature, transitional, mature, memory IgD(+), memory IgD(-), and plasmacytes isolated from the blood of a single individual. Our results support the view that in both human and mouse, the H chain repertoires expressed by individual, developmental B cell subsets appear to differ in sequence content. Sequencing of unsorted B cells from the blood is thus likely to yield an incomplete or compressed view of what is actually happening in the immune response of the individual. Our findings support the view that studies designed to correlate repertoire expression with diseases of immune function will likely require deep sequencing of B cells sorted by subset.
Keywords: B cells subsets; CDR-H3; human antibody repertoire.
Figures

References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources