

Evolutionary origin of gastrulation: insights from sponge development
- PMID: 24678663
- PMCID: PMC4021757
- DOI: 10.1186/1741-7007-12-26
Evolutionary origin of gastrulation: insights from sponge development
Abstract
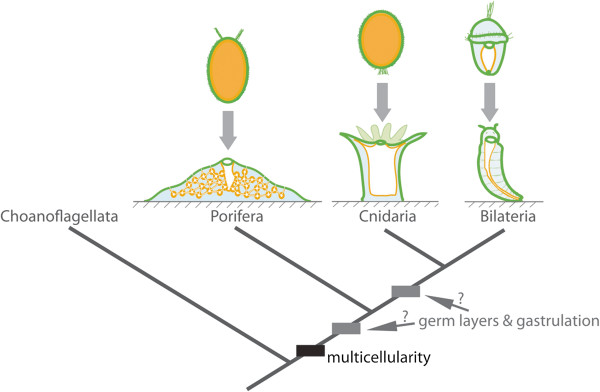
Background: The evolutionary origin of gastrulation--defined as a morphogenetic event that leads to the establishment of germ layers--remains a vexing question. Central to this debate is the evolutionary relationship between the cell layers of sponges (poriferans) and eumetazoan germ layers. Despite considerable attention, it remains unclear whether sponge cell layers undergo progressive fate determination akin to eumetazoan primary germ layer formation during gastrulation.
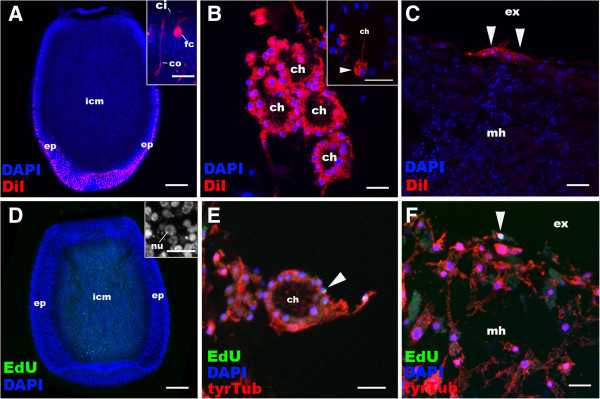
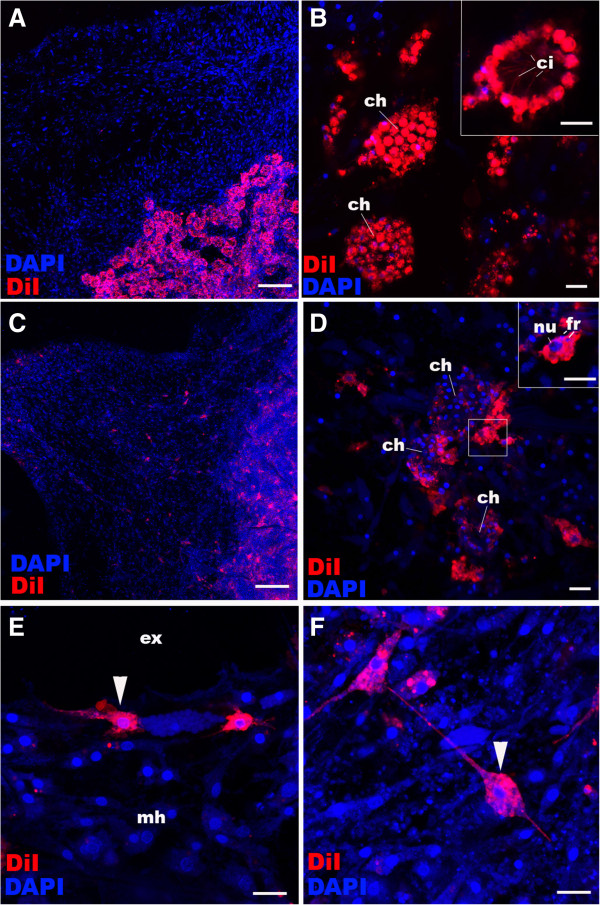
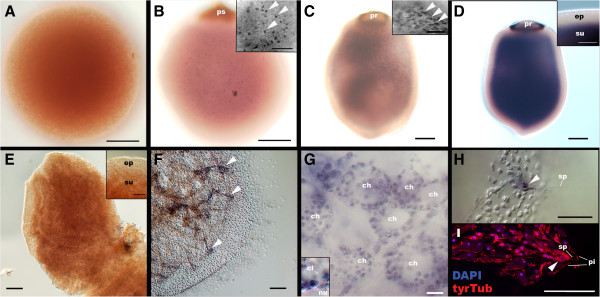
Results: Here we show by cell-labelling experiments in the demosponge Amphimedon queenslandica that the cell layers established during embryogenesis have no relationship to the cell layers of the juvenile. In addition, juvenile epithelial cells can transdifferentiate into a range of cell types and move between cell layers. Despite the apparent lack of cell layer and fate determination and stability in this sponge, the transcription factor GATA, a highly conserved eumetazoan endomesodermal marker, is expressed consistently in the inner layer of A. queenslandica larvae and juveniles.
Conclusions: Our results are compatible with sponge cell layers not undergoing progressive fate determination and thus not being homologous to eumetazoan germ layers. Nonetheless, the expression of GATA in the sponge inner cell layer suggests a shared ancestry with the eumetazoan endomesoderm, and that the ancestral role of GATA in specifying internalised cells may antedate the origin of germ layers. Together, these results support germ layers and gastrulation evolving early in eumetazoan evolution from pre-existing developmental programs used for the simple patterning of cells in the first multicellular animals.
Figures
References
-
- Hyman LH. The Invertebrates: Protozoa Through Ctenophora, Volume 1. McGraw-Hill Book Company, Inc: New York and London; 1940.
-
- Hall BK. Germ layers and the germ-layer theory revisited - primary and secondary germ layers, neural crest as a fourth germ layer, homology, and demise of the germ-layer theory. Evol Biol. 1998;12:121–186.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
